科技工作者之家
科界APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2020-03-23
来源:宏基因组
病原菌之外:微生物组与植物免疫系统的相互作用Beyond pathogens: microbiota interactions with the plant immune systemCurrent Opinion in Microbiology [6.916]
Volume 49, June 2019, Pages 7-17
原文链接:https://doi.org/10.1016/j.mib.2019.08.003
第一作者: Paulo José PL Teixeira
合作作者:Nicholas RColaianni, Connor RFitzpatrick
通讯作者:Jeffery L Dangl
主要单位:北卡罗来纳大学教堂山分校生物学系和霍华德·休斯医学院(Department of Biology and Howard Hughes Medical Institute, University of North Carolina at Chapel Hill, Chapel Hill, University of North Carolina at Chapel Hill, NC, United States)
划重点标题:病原菌之外:微生物组与植物免疫系统的相互作用
关键字:植物微生物组、植物免疫,免疫受体,MAMP,MTI,ETI
植物免疫受体可以接收微生物分子信号并触发一系列有效的生化反应,抵御大多数入侵者。
目前,植物免疫系统对于检测和控制致病菌的作用已被充分描述。
相比之下,人们对在复杂微生物背景下生存的植物,及其免疫的了解还很少。
近期研究表明,与致病菌类似,共生植物微生物组也可以抑制或逃避宿主免疫。
植物免疫系统对微生物组的组装具有积极作用,并通过响应环境变化控制微生物稳态。
植物免疫系统塑造了微生物组,并且微生物组可增加植物免疫能力,并充当抵抗致病菌的一层附加防线。
背景Introduction
自1980年代中期以来,科学家一直在研究植物免疫的分子机理。
关于植物如何识别,响应和限制入侵生物的生长,以及病原体用来抵抗植物免疫策略的研究,导致了植物免疫系统的概念框架的形成。简而言之,植物具有识别非自我分子或修饰自我分子的受体,这些分子表明潜在的入侵者的存在。位于质膜的第一层模式识别受体(pattern recognition receptors,PRR)可以感知细胞外分子的存在,这些分子通常在各种微生物类别中都较为保守(例如真菌的几丁质或细菌的鞭毛蛋白),因此这些被称为微生物相关分子模式(Microbe-Associated Molecular Patterns,MAMP)。宿主识别MAMPs触发的免疫应答称为MTI(MAMP-triggered immunity),这足以阻止大多数微生物的侵染。但是,一些具适应性的致病菌已经进化出效应分子来干扰MTI以及宿主的生理,细胞外的MAMPs和细胞内的效应因子的界限越来越模糊。反之,植物通过第二级受体来抵御产生抗病性的病原体。这些受体属于NLR蛋白家族(Nucleotide-binding Leucine-rich Repeat,核苷酸结合的富含亮氨酸的重复序列),充当细胞内传感器的功能,识别特定的效应蛋白。通过相应的特定宿主NLR蛋白直接或间接的感应病原菌效应子从而触发的免疫称为ETI(Effector-triggered immunity),这是一种强大的抗病反应,通常包括局部宿主细胞死亡并触发系统防御信号。控制防御的植物激素与控制植物生长的植物激素之间的复杂相互作用是植物免疫系统的重要组成部分。
这种模式很好地描述了植物免疫控制的基本原理,但是它基于植物和病原微生物的互作。然而,很明显,植物和多种共生微生物之间也有密切的关系,在很多环境下宿主植物地上和地下组织中都形成复杂的群落(如微生物组)。实际上,很多微生物和植物之间的互作关系是非致病的,但微生物中一些表达的分子可能被植物免疫系统识别。因此植物微生物组研究的一个主要问题就是植物是否能识别,或怎样区分共生微生物和病原体。病原微生物规避植物免疫的策略是否适用于微生物组?微生物组可以促进植物免疫吗?环境的变化如何影响植物免疫和微生物组之间的相互关系?这些是新兴领域中的一些关键问题,近期已得到越来越多的关注。本文中,我们回顾了与植物免疫系统和微生物之间的相互作用相关的最新研究和新颖概念,主要侧重于拟南芥及其相关物种作为研究模型的实用性。
正文植物免疫系统影响微生物组装配Evidence for the participation of the plant immune system in microbiome assembly
宿主植物微生物群落组成受周围环境影响,但控制这些微生物装配的机制还依然未知。共生假说认为植物及其微生物组受进化过程影响形成一个整体,这意味着植物已经采用了一种适应性方法,将其”进化伴侣”与其他微生物分开。另外一种说法是,一些微生物组的组装可能代表了生态位补充,该过程受到植物性状影响,但是对植物宿主的适应性重要性影响较小。不管目前提出的假说对于微生物组整体的适应性如何,很明显共生群落的某些部分对宿主具有适应性价值。通常,普遍认为植物可以区分致病菌和共生微生物。实际上,在对密切相关的共生和致病假单胞菌的分析中,极少数的毒力因子的产生或缺失直接导致了生存方式之间的转换。此外,通过对拟南芥根系和叶片627个细菌基因组的检测,发现608个细菌(占97%)有可能产生免疫原基因MAMPs(图1a)。这些细菌中许多具有与已知病原体共有相同MAMP变体,表明抑制病原菌生长的MTI反应也可能响应并被激活,这就引出了一个问题,即共生菌如何避免和抑制MTI。MTI可以通过与宿主的亲密接触而从内而外的作为门控来抑制病原体并维持微生物组的稳态。这种观点认为,MTI是植物控制微生物群落装配的一种机制。研究表明,在MAMP识别和下游信号MTI转导方面有缺陷的拟南芥多突变体,对正常的致病丁香假单胞菌无毒突变体的防御能力降低,并且无法在高湿度下维持叶片正常的内生细菌群落,无法调节内生细菌群落的生长导致某些叶片出现萎黄和坏死,类似营养不良的症状。这表明植物通过其免疫系统控制微生物组成员的生长,以维持自身健康。
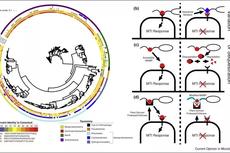
图1. MAMP的常规模式和微生物用来逃避植物免疫系统的潜在机制Figure 1. Commensal MAMP profiles and potential mechanism used by microorganisms to evade the plant immune system.
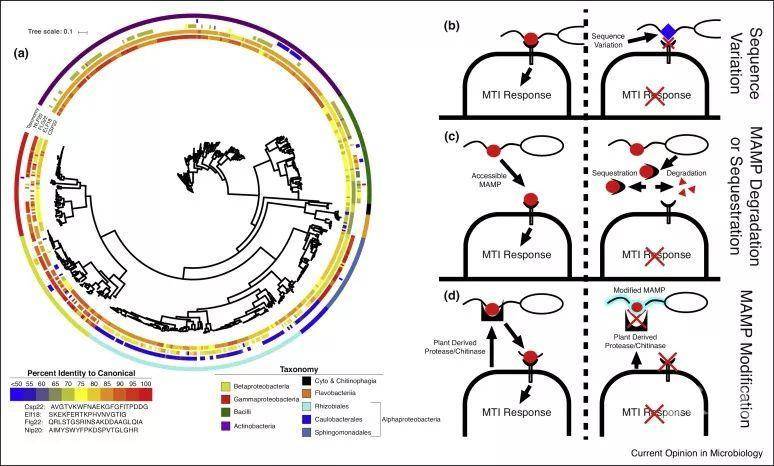
(a) 从健康的拟南芥植物中分离到的细菌具有潜在免疫原性MAMP,而其他细菌则和标准序列存在很大差异,这可能有助于共生菌逃避植物免疫系统。图中包括627个分离自拟南芥的菌株,Levy等人生成3837个微生物的树。MAMP基因通过定制的隐型Markov模型进行鉴定,并将所有MAMP基因和MUSCLE比对后进行鉴定,使用编辑距离(生成)计算同一性百分比。该树是使用iTOL生成的,标准序列来源于Micrococcus lysodeikticus (csp22) , Escherichia coli (elf18), Phytophthora parasitica (nlp20) 以及Pseudomonas syringae pv tabaci (flg22)。MAMP的差异导致逃避植物免疫系统可能通过(b) 序列变异、(c) MAMP降解和螯合、(d) MAMP修饰等。详见正文描述。
微生物组用于逃避或抑制植物免疫的策略Strategies used by the microbiome to evade or suppress plant immunity
植物免疫受体不能区分微生物的生存方式,并且识别在病原菌和共生菌中都存在的受体。逃避和抑制宿主免疫反应是成功的致病菌的标志。同样地,植物微生物组个体成员的定殖,即群落组装的本质,可能都需要采取相应的策略来逃避或者干扰植物免疫。最近的工作凸显了用于抑制和逃避植物免疫系统的策略中,致病菌和共生菌之间的异同。
病原菌利用抑制MTI来逃避植物免疫系统,但最近也有非致病微生物的相关报道。有益的根际细菌假单胞菌WCS417促进植物生长并抑制由细菌MAMP flg22触发的部分转录反应。同样地,在拟南芥根系定殖的特定根瘤菌能够阻止由同一MAMP触发的反应。内生菌还可以防止由MAMP出发的拟南芥中胞质钙离子内流。最近的研究表明,促进植物生长的细菌WSC358产生的有机酸会降低细胞外pH,并干扰flg22的反应。另外,通过抑制植物先天免疫,共生真菌可以进入植物组织。然而,由共生菌产生的对免疫反应机制的抑制亟待研究。III型分泌系统(T3SS)是致病细菌中的一个共同特征,在非致病菌中也有发现。但是编码这种效应子-传递机制的基因在植物相关共生细菌的基因组中还很少见。这可能反映了大多数与植物相关的共生菌在宿主特异性较低。因此,在植物微生物组研究中有多种干扰素住免疫反应的替代策略,尤其是与细胞外MTI相关的反应。
逃避MTI是病原微生物和共生微生物侵染植物的另一种策略。微生物已进化出至少三种逃避MTI的机制:
(1)MAMP序列变异,
(2)MAMP降解/螯合和
(3)MAMP修饰。
MAMP序列变异MAMP divergence
微生物通过进化为MAMP的变种,不再结合或激活相应的植物PRR。由于MAMP序列和结构的改变可能会损害微生物的含MAMP分子的有益功能,这种逃避可能在表面上的产生反适应性(counter-adaptive)。例如,一些失去免疫原性的flg22变体也失去活力。然而,在某些细菌类群中广泛存在多种可能逃避免疫的MAMP变体。这种现象可能是依赖MAMP的(图1a):在拟南芥相关细菌分离物中发现的26%的flg22肽表位与至少50%的标准活性序列存在差异,而鉴定出的elf18变体中只有不到1%与标准表位的差异至少为50%。这表明flg22识别使细菌适应性缺陷变强,和/或flg22区域相比elf18更易于变异,和MAMP识别的要求不同。与第一个假设一致,elf18受体EFR不在拟南芥根中表达,而FLS2则在根系表达。与相应的标准序列不同,MAMP序列产生的MTI响应依然未知。与在与植物相关的细菌中发现,A,P序列多样性的同时,拟南芥和番茄品系对不同MAMP甚至对相同MAMP变体的反应也表现出很大的差异。这表明植物群体对MAMP的认识也正在发展中。因此这种识别可能是由宿主本身共生和有益微生物的MAMP所驱动的,不仅仅是病原体。前者的抗性和后者抗性必须相对平衡。
MAMP降解和隔离MAMP degradation/sequestration
即使微生物表达具有免疫原性的MAMP,它们可能也具有逃避MTI的机制。一些微生物已经进化出可以消化其MAMP的蛋白酶,或者可以将MAMP或蛋白质与植物受体隔离(图1b)。植物病原体丁香假单胞菌DC3000分泌一种蛋白酶,该酶通过鞭毛蛋白的降解降低MTI并增加该菌株在拟南芥和番茄叶片中的生长。几丁质是真菌中细胞壁的一种保守成分,是另一种有效的MTI诱导剂。几种真菌病原体已经进化出几丁质结合蛋白(LysM或失活的几丁质酶),它们能够隔离游离的几丁质,从而阻止植物PRR的激活。尽管,迄今为止仅在病原体中证明了这种机制,但在整个真菌界中都发现了LysM和可能无活性的几丁质酶,因此,共生菌可能使用这种方法逃逸MAMP。
MAMP修饰MAMP modification
微生物阻止MTI触发的另一种策略是MAMP修饰(图1c)。例如,本氏烟草(Nicotiana benthamiana)分泌糖苷酶,该糖苷酶从细菌鞭毛中剥去了聚糖屏蔽,使植物蛋白酶释放出免疫原性的flg22肽,供FLS2受体识别。反过来,病原体可以通过抑制植物糖苷酶或修饰覆盖其鞭毛的聚糖部分来逃避flg22的识别。同样,真菌可以通过将细胞壁中的几丁质脱乙酰降解为壳聚糖来逃避植物免疫系统(壳聚糖是一种较弱的免疫诱导剂)。因为flg22和几丁质都在植物微生物组中普遍存在,所以共生菌很可能进化出类似的MAMP修饰策略来逃避MTI。
图2. 微生物组扩展了植物免疫系统Figure 2. The microbiome expands the plant immune system
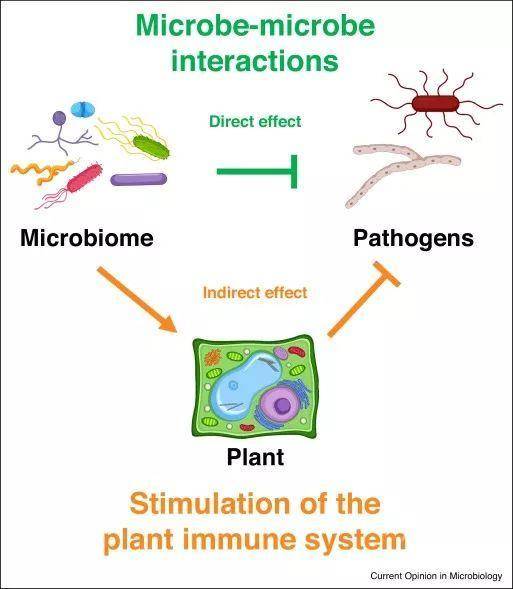
植物微生物组可以通过至少两种机制充当对于致病菌的额外防御层。首先,微生物组可以通过产生抗菌化合物或通过生态位竞争直接抑制致病菌的定殖;其次微生物组可以通过刺激植物免疫系统间接提高对病原菌的抵抗能力,从而增强抗病性。
微生物组是植物免疫系统的延伸The microbiome functions as an extension of the plant immune system
一般对植物病害的研究通常关注宿主和病原体之间的双元互作。然而,近期研究表明微生物组可以扩展植物的防御能力,并且常常影响植物和病原体之间的互作的结果,并且通过包括“生物防治”等未知机制来预防或减轻疾病的发生。重要的是,到目前为止,这似乎很大程度上仅仅由两个主要机制决定:一个是微生物的直接相互作用;另一个是刺激或引发植物免疫(图2)。
微生物之间的相互作用在抑制病原体方面起着越来越明显的作用,可以作为抵御植物中入侵生物的第一道防线。例如假单胞菌ZJU60株分泌的分子是从被感染的小麦穗部分离的,它通过抑制一种组蛋白乙酰转移酶抑制禾本科镰刀菌生长。此外,最近的一项综合研究表明,从拟南芥叶片微生物组中分离出来的细菌的共同特征,就是与其他微生物(包括致病菌)产生拮抗作用。进一步挖掘基因组揭示了具有抑制性的菌株中高度流行的各种未知生物合成基因簇,并鉴定了两种新型抗生素。研究表明,植物微生物组是可以直接抑制病原体拮抗剂的丰富来源。另外还有很多其他因素有助于微生物组直接抑制病原菌,有研究提出微生物组与入侵致病菌形成资源竞争和生态位重叠,从而形成限制植物病原体入侵的重要因素。这类似于共生微生物保护动物肠道,在该动物肠道中,入侵的有害微生物无法与之竞争,宿主微生物组抑制了它们的生长。值得注意的是,拟南芥根系天然潜伏着有害丝状真菌,但在多种微生物组的背景下,这些真菌仍受到多种共生细菌的控制。如果去掉这些保护性细菌会导致植物致病,因此,微生物组稳态和微生物之间的互作对植物健康至关重要。更多信息请阅读 《Cell :根部微生物跨界的互作促进拟南芥生存》
除了上述对致病菌的直接抑制外,共生微生物还可以通过刺激植物免疫系统来促进宿主健康,从而间接地抑制疾病。植物中微生物组介导的免疫的一种众所周知的形式,即诱导的系统抗性(ISR),它是针对由根相关微生物触发的叶片病原菌和害虫的防御反应。已有研究报道了ISR所需的宿主遗传成分,许多与系统发育无关的微生物在许多不同的植物物种中可引发ISR。ISR的标志是宿主敏感性增强,而不是防御基因的组成性激活。这意味着ISR可促进更快更强的全身免疫反应,但仅在受到刺激时才如此。还有研究报道了微生物组介导的抗病性,涉及植物免疫系统的组成性激活(警报状态)。从拟南芥中分离出的一株鞘氨醇单胞菌可激活部分植物防御基因,并增强针对细菌病原体丁香假单胞菌DC3000的免疫力。这种保护作用在bak1/bkk1突变体中消失,表明这种共鸣可能依赖于BAK1作为共受体的PRR通过MAMP识别来刺激植物免疫。这些研究表明,通过共生微生物抑制疾病可能需要完整的植物免疫系统,这进一步证明微生物组参与植物与病原体的相互作用。
近期研究表明植物微生物组都是动态变化的,并且会对致病菌和害虫的存在做出响应,这支持了一个有趣的假设,即植物在某些条件下会主动选择共生微生物来保护自己抗击疾病。荷兰田间自然土壤中生长的拟南芥中,霜霉病病原菌Hyaloperonospora arabidopsidis感染拟南芥叶片之后,促进了根际三种特定的诱导ISR的细菌增殖,并且这些保护细菌似乎在土壤中持续存在,并为随后植物起到抗霜霉病的作用。丁香假单胞菌侵染拟南芥叶片也会导致类似的土壤菌群变化,从而保护后代植物。为了支持宿主在重塑微生物菌群过程中的积极作用,被侵染的植物显示出根系分泌物渗出等现象,推测有利于选择有益细菌。在侵染F. graminearum的大麦根系,以及被粉虱侵染的辣椒幼苗中,也观察到了微生物组成的变化和潜在有益微生物的富集。重要的是,根际中有益微生物的富集与疾病抑制土壤的发展有关,在土壤中即使存在致病菌,植物也能保持健康。因此,微生物组的改变可能影响在同一土壤中种植的植物的后代,从而影响生态和农业进程。
可进一步阅读本组参与发表的抑病土壤相关文章 - Microbiome:根系分泌物驱动土壤记忆抵御植物病原菌。
与ISR形成鲜明对比的是,一些与根相关的细菌可使宿主植物更易感染叶片致病菌。在拟南芥中增强对草食动物抗性的假单胞菌菌株,还能引起对半生营养性致病菌的诱导全身敏感性(induced systemic susceptibility,ISS)。这涉及到细菌产生的亚精胺,但该分子如何调节植物免疫力尚不清楚。此外,丛枝菌根真菌还增加了豆科植物黄芪冲剂对叶面病原体Erysiphe pisi的敏感性,这会引起白粉病。这些研究表明,共生微生物可以对植物的免疫系统产生多效作用,表明在农业中有效地通过免疫调节微生物,取决于微生物组对植物以及致病菌或其他共生微生物在不同的环境中的不同影响。
图3. 包括温度,水或养分利用率的差异在内的环境变化,可以通过多种机制调节植物的免疫力Figure 3. Environmental variation, including differences in temperature, water, or nutrient availability can modulate plant immunity by a number of mechanisms.
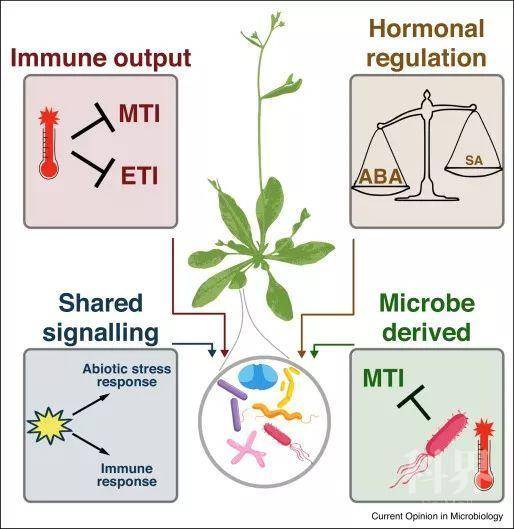
首先,不同的环境可以影响植物免疫产物的表达,包括MAMP的成分和效应器触发的免疫。其次,非生物胁迫激素(如脱落酸)和防御激素(如水杨酸)之间的相互影响可以调节植物免疫。第三,共同的信号传导成分可以导致植物联合非生物胁迫和免疫反应。最后,微生物可以在特定环境下调节植物的免疫力。植物免疫调节的所有这些机制都有改变植物微生物组的潜力。
环境相关的免疫调节过程可以塑造环境中的植物微生物组Context-dependent immune modulation shapes the plant microbiome across environments
自从在陆地环境中定居以来,植物就面临着温度,水和养分利用率以及盐度,pH和重金属等化学属性不断变化的环境。植物对环境变化的响应与植物免疫相互关联,部分原因是植物需要对非生物胁迫和致病菌入侵做出最恰当的响应。然而,这种相互联系对植物微生物组的组装和功能具有一定的影响,目前还在研究中。
环境变化可以通过许多非排他性机制来调节植物免疫(图3)。首先,不同环境可以直接调节植物免疫输出的表达。例如,在环境胁迫下植物会调节MTI和ETI相关基因的表达,而当温度升高时,一些NLR受体会失去功能。第二,植物响应非生物胁迫会调节激素水平,如抑制茉莉酸和水杨酸的防御途径而对植物免疫产生拮抗作用。这种作用非常大,以至于一些致病菌切断这种互作从而使其能够入侵植物。近期研究表明,非生物响应对植物免疫力的拮抗作用取决于多种因素,例如植物年龄和胁迫因子先后顺序,最后,共有的信号传导成分可以共同协调植物对非生物胁迫和植物免疫的反应。例如,PRR几丁质激发子受体激酶1(CERK1)不仅可以增强植物对真菌病原体的防御能力,而且还可以强烈调节盐胁迫所需基因的表达。
微生物也可以调节不同环境下的植物免疫。在较高温度下,细菌效应子向宿主的转运增加,这有助于抑制植物的免疫。此外,当寄主植物在低磷酸盐条件下生长时,内生真菌Colletotrichum tofieldiae的定殖会降低植物免疫。变形杆菌属的成员会在营养耗尽的情况下排出其极鞭毛,这可能是为了在不利条件下节省能量。但是,由于鞭毛蛋白是植物免疫力的重要触发因素,在特定环境下鞭毛的丧失也可能导致MTI的调节,并对随后的微生物定殖产生潜在的后果。有趣的是,内共生弧菌 Vibrio fischeri 在夏威夷短尾鱿鱼的某器官中的持续存在,并伴随鞭毛的丧失,这表明通过MAMP修饰对宿主免疫力的环境依赖性调节,可能是动植物宿主的相关微生物组的一个共同的特征。
在环境或营养胁迫条件下调节植物免疫力可以重塑植物菌群。在低磷酸盐条件下,募集内生真菌C. tofieldiae需要植物免疫相关化合物,尤其是色氨酸衍生的次生代谢产物。另外,植物磷酸盐饥饿反应的主要转录调节因子PHR1通过靶向SA和JA途径中的基因直接调节植物免疫力。这种互作会减弱植物的免疫反应并扰乱根系微生物组。拟南芥在铁饥饿或有ISR诱导细菌的情况下产生的根系分泌物香豆素,在根际中可发挥选择性抗菌作用,抑制真菌病原体并影响微生物组群落结构,选择有益的微生物响应铁饥饿,植物合成香豆素并将其渗入根际,在有助于铁在土壤中转移的同时,还通过其抗菌活性重塑根部微生物组的组成。PBS3以非依赖于年龄的方式调节非生物胁迫下的植物免疫,也重塑了叶片细菌群落的组成。
关于香豆素研究,可进一步阅读本组参与的研究:PNAS:植物香豆素塑造拟南芥合成根系微生物组的组成。
尽管越来越多的证据支持植物免疫调节可以重塑植物微生物群的想法,但这种微生物组重塑对植物性能的影响,无论是正面的还是负面的,仍需探索。植物在低铁条件下渗出的香豆素选择性抑制病原真菌的生长,同时维持促进植物生长的假单胞菌和其他微生物的生长。这些香豆素选择的微生物可以通过增加植物对铁饥饿的耐受性来使植物受益。同样,在食草动物,干旱和致病菌侵袭等胁迫下,植物可能会受益于其微生物组的变化。尽管在这些例子中,微生物组改变的潜在机制尚未确定。在低磷酸盐条件下,真菌内生真菌C. tofieldiae的募集使拟南芥植物的生物量增加。然而,人工重组菌群实验表明,在磷饥饿下,富集拟南芥根细菌分类群抑制植株生长,这是通过茎磷酸盐的积累来衡量的。最近研究表明,拟南芥产生的三萜是根细菌微生物组的主要决定因素。这些分子根据细菌类群起抗生素或增殖剂的作用,并选择性地调节根微生物群的组成。重要的是,可以响应非生物胁迫以及在病原体和草食动物侵袭期间诱导三萜生物合成。这些例子表明,部分环境由植物免疫调节介导的整个环境中微生物组的改变对植物的生长既有正面影响,也有负面影响。
关于三萜和二半萜对微生物组的影响,请阅读:
Science:拟南芥根系三萜化合物塑造特异的微生物组
SCLS:拟南芥二半萜类化合物调控根系微生物组
结论Conclusions
我们对植物免疫系统的了解主要基于数十年来对植物与致病菌相互作用的研究。现在正在微生物学的背景下重新研究,测试和构建这种知识,揭示出令人兴奋的差异和相似之处。迄今为止的研究表明,植物免疫系统同样介导与病原微生物和共生微生物的相互作用。被植物免疫受体识别的大多数MAMP通常在病原体和共生体中发现,这表明微生物对植物组织的定殖涉及抑制、逃避宿主免疫系统,无论其生活方式如何。实际上,最近的工作表明许多共生细菌可以抑制MTI。未来的研究应关注这种抑制的机制及其对微生物组组装的意义。同样重要的是病原体与共生微生物之间的相互作用。我们认为微生物组可作为植物免疫系统的额外附加功能,可以通过微生物与微生物的相互作用直接抑制疾病,也可以通过刺激植物免疫来间接抑制疾病。我们预测,病原体必须具有进化的策略来克服微生物组所施加的免疫屏障。最后,植物的免疫系统与环境和养分反应有着复杂的联系,面对环境变化,微生物群落的变化常常可见。一个相关的方向是研究环境相关的免疫调节如何改变植物微生物群的组成,以及如何帮助植物在各种环境下的胁迫条件下的成长。确定植物免疫力和微生物组之间复杂相互作用的潜在机制和由此产生的植物响应,是推进植物与微生物相互作用领域的重要目标。
来源:meta-genome 宏基因组
原文链接:https://mp.weixin.qq.com/s?__biz=MzUzMjA4Njc1MA==&mid=2247489735&idx=2&sn=703baf6b8a2f56bde9cd73d9b0f11a27&chksm=fab9f276cdce7b60b1c3afe209ad2829c9e4a83709e5ad43cc4f3bab6efba21cf055d9b5329c#rd
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn
微生物学: 生命早期的抗生素使用可能会影响发育

研究揭示拟南芥响应病原体信号快速诱导植保素合成的机制!
疾病: 了解风湿性关节炎的发病机制
阴道病原体检查
阴道病原体检查
美国院士:强烈呼吁中国母亲不要给小孩滥用抗生素


COM:病原体之外-微生物组与植物免疫系统的相互作用
脑脊液病原体检查(CSF)
无特定病原体级实验动物
浆膜腔积液病原体


科界APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号