

科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2020-11-17
基因剪接是通过一些酶学操作使一条DNA分子与另一条DNA分子相连。即在mRNA成熟期,切除基因的内含子,连接基因的外显子的过程,称为基因剪接。而天然基因的某些片段被合成的DNA链所取代或连成整体的过程称为基因剪辑。一个基因为它的等位基因所替换,而其他基因则保持不变称为基因置换1。
简介基因组中或基因组间发生遗传信息的重新组合,被称为DNA重组(DNA recombination),其中发生在基因组中的DNA重组又称DNA重排。包括同源重组、特异位点重组和转座重组等类型,广泛存在于各类生物。体外通过人工DNA重组可获得重组体DNA,是基因工程中的关键步骤。
类型同源重组同源重组( homologous recombination)是指发生在两段同源序列之间的DNA片段交换。两段同源序列既可以完全相同,也可以存在差异,既可以位于两个DNA分子上,也可以位于一个DNA分子中。真核生物的同源染色体交换及姐妹染色单体交换、细菌的转导和转化、噬菌体的重组都属于同源重组。有多种模型阐述同源重组机制,这里介绍其中两种。
(一)Holliday模型
Holliday于1964年提m Holliday模型,将同源重组分为四个阶段。
1.同源序列配对。
2.形成Holliday结构,即两段同源序列的单股同源DNA的同一磷酸二酯键被水解,同源末端交换,连接,形成Holliday结构(HoIJiday structure,又称Holliday连接体)。
3.形成异源双链DNA,即Hollidav结构发生分支迁移,形成异源双链DNA(hetero-duplex DNA。
4.Holliday结构解离,即两段同源序列的单股同源DNA的同一磷酸二酯键被水解,Holliclay结构解离,连接切口,形成重组体。水解位点不同,所得到的重组体也就不同:
(1)两次水解的是同股DNA,形成片段重组体(patch recombinant)。这种重组未发生实质性交换,依然是A-B、a-b。
(2)两次水解的是异股DNA,形成拼接重组体(splice recombinant)。这种重组发生了实质性交换,产物是A-b、a-B。
大肠杆菌Holliday结构的解离由解离酶(resolvase) RuvC催化。RuvC是同二聚体,每个亚基含172AA。
(二)双股断裂修复模型
双股断裂修复模型( double-strand break repaii。mnodel)也将同源重组分为四个阶段(图2-32)。
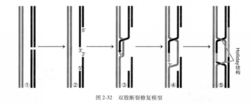
1.同源序列配对。
2.形成3’端突出结构,即配对同源序列之一的DNA双链水解,并由5’外切核酸酶水解,形成3'端突出结构(即3’黏端)(①~②)
3.形成HollidaY结构,即一个3’端攻击另一段完整的同源序列,随后发生分支迁移,形成Hollidav结构(③~⑤)。
4.HollidaV结构解离,两种解离方式得到两种不同的重组结果。
位点特异重组位点特异性重组(sile-specific recombinalion)是指发生在特定的DNA序列之间的片段交换,序列之间不要求有同源性。位点特异性重组发生于包括基因表达调控、胚胎发育过程中的程序性基因重排、免疫球蛋白基因的重排、一些病毒DNA和质粒复制周期中发生的整合与解离等过程中。
(一)重组机制
位点特异性重组本质上是两个重组位点的四股DNA发生两次切割和两次连接的过程,所需的关键成分是重组酶( rccombinase),此外还需要一些蛋白因子。这里以入噬菌体DNA与大肠杆菌DNA整合而进入溶原状态为例,介绍位点特异性重组机制(图2-33)。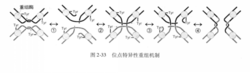
1.第一次切割重组酶(又称入噬菌体整合酶,integrase,356AA,由入噬菌体基因组编码)四聚体与两个重组位点(入噬菌体重组位点attP和大肠杆菌重组位点au,B有15bp相同的核心序列)结合,切断两个重组位点一股DNA特定的磷酸二酯键,形成两个切口,切口3’端的磷酸基与重组酶活性中心Tyr342的羟基以磷酸酯键结合。
2.第一次连接两个切口的5 7端交换,与对方3’端连接,形成HoUiday结构。
3.第二次切割重组酶切断两个重组位点另一股DNA特定的磷酸二酯键,形成两个切口。切口3 7端的磷酸基与重组酶活性中心Tyr342的羟基以磷酸酯键结合。
4.第二次连接两个切口的5’端交换,与对方3’端连接,Holliday结构解离。
有些位点特异性重组体系的两个重组位点的四股DNA可能同时切割,同时连接,并不形成Holli-day结构。
(二)重组效应
位点特异性重组既可以发生在一个DNA分子中,也可以发生在两个DNA分子间。重组酶识别位点有方向性,所以重组时两个重组位点的排列有方向性。
1.插入 当位点特异性重组发生在两个闭环DNA之间或一个闭环DNA与一个线性DNA之间时,重组的结果是DNA插入(即整合),并且插入之后在两端形成同向重复序列(DR,图2-34①从下向上)。
2.缺失 如果DNA分子中一个片段的两端存在同向重复序列,则该DNA分子可以通过位点特异性重组使该片段缺失,并且缺失片段成环(图2-34①从上向下)。
3.倒位如果DNA分子中一个片段的两端存在反向重复序列(1R),则陔DNA分子可以通过位点特异性重组使该片段倒位(图2-34②)。
转座重组1944年,McClintock(1983年诺贝尔生理学或医学奖获得者)在研究玉米基因时发现:有些DNA片段可以在染色体DNA中移动位置。现已阐明:基因组DNA巾存在一些非游离的、能自复制或自剪切、并能以相同或不同拷贝在基因组中或基因组间移动位置的功能性片段,被称为转座元件( transposableeleInent)。转座元件或其拷贝移动位置的现象称为转座( transposition)。大多数DNA转座对转座位点的选择既不要求有同源性,也不要求有特异性,几乎是随机的。
转座元件有很多种类,例如以下两类:①转座子(Tn):最初称为跳跃基因( jumping gene),具有完整转座元件的功能特征,可携带内外源基因组片段(单基因或多基因),可表达出新的表型,由白己编码的转座酶(transposase)催化转座。②逆转录转座子又称逆转录子( retrotransposon,retroposon):通过转录、逆转录进行转座的转座元件。这里介绍转座子及其转座。
(一)转座子
转座子在原核生物和真核生物中普遍存在,、细菌有两类典型的转座子:简单转座子和复合转座子。
1.简单转座子( simple transposon) 又称插入序列(IS),是结构最简单的转座子,长度为700~1531bp,由转座酶基因序列和两端长度为9~41bp的反向重复序列构成,其中反向重复序列是转座酶识别位点(图2-35) 。
。
2.复合转座子( composite transposon) 除了含转座酶基因序列和反向重复序列之外,还含与转座功能无关的其他基因序列,这些基因常常赋予宿主细胞某种表型,例如抗性基因赋予宿主细胞抗药性。复合转座子可以进一步分类,例如Tn3转座子就是一种含氨苄青霉素抗性基因的复合转座子。
(二)转座机制
细菌转座子有两种转座机制:简单转座和复制转座。
1.简单转座( simple transposilion) 转座酶将转座子从原位点切下,插入被转座酶错位切割的转座位点,经过填补之后,两端形成短的同向重复序列(4~13bp)。原位点或被连接修复,或所属DNA被降解。降解通常是致死性的(图2-35)。
2.复制转座( replic:ative transposition) 在原位点与转座位点之间形成共整合体,包括以下步骤:①转座酶切割转座子两端,形成切口。②转座子的两个3’.羟基错位攻击转座位点的两个3’,5’一磷酸二酯键。③转座子与转座位点共价连接,转座子复制,形成共整合体(cointegrate,又称共合体)。④共整合体重组解离,原位点与转座位点各有一个转座子(图2-36)。
(三)转座效应
DNA转座可以影响转座位点基因的功能和活性:①转座位点位于编码序列内,转座子插入导致基因突变。②转座位点位于调控序列内,转座子插入影响基因表达。③在转座位点插入转座子基因,赋予新表型,例如抗药性。④链内复制转座后,转座子拷贝之间发生位点特异性重组,导致缺失或倒位2。
意义①参与DNA复制。
②参与DNA修复。
③参与基因表达调控。
④在真核细胞分裂时促进染色体正确分离。
⑤维持遗传多样性。
⑥在胚胎发育过程中实现程序性基因重排2。
历史1972年,加州大学旧金山分校的微生物学家赫伯特·伯耶(Herbert Boyer)、斯坦福大学的研究员史坦利·科恩(Stanley Cohen)在火奴鲁鲁参加学术会议时在一家现成食品店里遇到了对方。他们一边吃着熏牛肉三明治,一边构思除了一个开创了现代生物技术产业的实验。回到加州后,这两个人成功的将青蛙的DNA插入到细菌中,这些细菌开始自我复制,同时也复制了插入其中的青蛙基因。这种技术被称为重组DNA,不过记者更喜欢称之为基因剪接3。
本词条内容贡献者为:
张磊 - 副教授 - 重庆师范大学
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号