科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2019-12-27
来源:BioArt植物
原文 | Joan W. Conaway
编译 | 咸姐
责编 | 兮
今年是杰出科学家Robert G. Roeder教授和Pierre Chambon教授发现这3种活性真核生物RNA聚合酶的50周年纪念年,这一里程碑式的成就开创了真核生物转录的生物化学研究的黄金时代,在此期间,随着技术的发展,人们对RNA聚合酶的结构和功能的研究有了巨大进展,同时对真核生物转录的机制也有了更多的了解。
真核生物中具有三种RNA聚合酶,即RNA聚合酶Ⅰ、Ⅱ、Ⅲ,它们分别参与不同类型基因的转录,对于基因的表达起着重要的作用,其中RNA聚合酶Ⅰ主要负责转录rRNA;RNA聚合酶II则转录mRNA以及一些小核RNA;RNA聚合酶Ⅲ负责转录5S rRNA、tRNA以及U6小核RNA等【1】。近20年,相比于RNA聚合酶Ⅰ和Ⅲ的转录过程,研究者们对RNA聚合酶II转录的起始与延伸过程的研究取得了更大进展。
RNA聚合酶介导的转录过程包括前起始复合物的形成、起始、延伸和终止。与原核生物RNA聚合酶直接识别基因启动子起始转录不同,真核RNA聚合酶不能直接识别启动子,需要形成前起始复合物,并在一系列转录起始因子的帮助下才能启动转录。
RNA聚合酶II(Pol II)的转录起始需要多种通用转录因子,包括TFIIB、TFIID、TFIIE、TFIIF和TFIIH【2】,这些转录因子是使酶能够从启动子精确启动所必需的,因此在最初发现和纯化Pol II之后的十几年里,大多数注意力都集中在识别和表征这些通用转录因子的努力上,然而随着研究的进展,有充分的理由表明Pol II介导的转录延伸调控也和起始调控一样复杂和有趣。
为了让我们能更好的了解这黄金50年里科学家们对于真核生物RNA聚合酶的相关研究所作的努力,近日,来自美国密苏里州斯托瓦斯医学研究所的Joan W. Conaway教授在Nature Structural & Molecular Biology上在线发表题为 The hunt for RNA polymerase II elongation factors: a historical perspective 的文章,结合包括自己在内的多个实验室的研究成果,详述了真核生物RNA聚合酶II(Pol II)延伸因子的发现史,并描述了目前从生物化学的角度定义在转录延伸过程中调控Pol II的转录因子的作用的性质和机制方面所做的一些重要工作。

生化、遗传和分子遗传学研究方法的结合最终确定了Pol II影响转录延伸过程的大量活动,其大致分为两类:第一类包括一系列直接针对Pol II延伸复合物并调节其活性的转录因子;第二类包括多种染色质修饰和染色质重塑酶,以及组蛋白伴侣,如SPT6和FACT复合物,它们可以通过染色质促进Pol II介导的延伸。而本文的作者将重点放在第一类延伸因子上,对其发现历史和研究工作做了详细综述。
对于Pol II延伸因子的探索始于上世纪七十年代,而第一个明显的成功是在东京大学的Shunji Natori实验室发现的Pol II延伸因子SII,现在也被称为“TFIIS”,它不仅是第一个被定义的Pol II延伸因子,其实也是第一个Pol II转录因子。随后十几年的研究逐渐形成了现在的SII作用模型(图1),按照这个模型,延伸酶既可以将下一个核苷酸添加到新的转录本中并进行正向移位,从而正确地定位Pol II活性位点中新添加的核苷酸的3’末端,也可以在DNA上向后滑动(“回溯”),从而导致转录本3’末端与Pol II活性位点的错配引起转录暂停或中止,而此时SII激活的转录本切割产生的新的3’末端将正确地定位在活性位点,继而重新开始延伸。
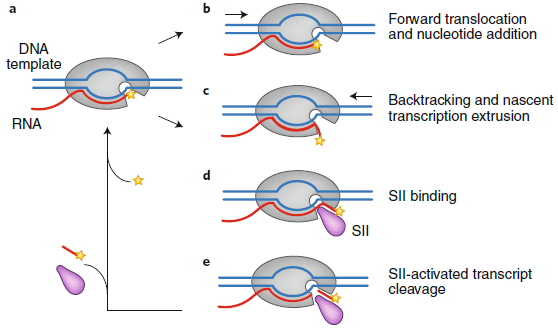
图1 中止Pol II依赖于SII的重激活
随后,研究者们又相继发现了第二类Pol II延伸因子,不同于SII的重激活作用,这一类延伸因子是通过抑制瞬时暂停和降低Pol II中止的频率来调控转录延伸速率的,包括Elongin、ELL-EAF复合物和TFIIF。
然而,正如让人印象深刻的故事总是充满情节反转的桥段一样,最初对于Elongin的研究表明其主要是作为转录的正向调节因子存在的,而随着研究深入,它的故事发生了戏剧化的转变,人们发现它不仅可以作为基因激活所需的转录因子,它也可以作为Cullin-RING泛素连接酶的底物识别亚基,有助于促进DNA损伤或其他情况诱导的停止Pol II的泛素化降解。
另一个意想不到的反转发生在对ELL-EAF的研究过程中。上世纪80年代末90年代初,研究人员发现蛋白酶抑制剂DRB阻断了Pol II诱导的延伸,从而激起了对DRB敏感的可调节Pol II介导的转录的方式的探索,继而发现了正向作用的DRB敏感的延伸因子——P-TEFb,研究表明DRB敏感的正转录延伸因子需要克服负转录延伸因子的活性,从而在转录起始不久后诱导Pol II的暂停,并伴随多个底物(包括Pol II CTD、NELF和DSIF)的磷酸化。进一步的,P-TEFb激酶被发现可以与ELL-EAF形成“超级延伸复合物(SEC)”,其既可以促进Pol II启动子近端停顿的释放,也可以在合成大量Pol II生成的转录本(包括编码热休克和其他应激反应分子的基因、c-Myc和其他肿瘤蛋白、HOX和其他发育调节因子以及增强RNA)时促进有效延伸。几乎在发现SEC的同时,有研究团队发现了第二个包含ELL-EAF的复合物,称为“小延伸复合物(LEC)”,尽管其详细功能还有很多未知,但LEC被证实可以影响转录本的起始和延伸。
综上所述,真核生物RNA聚合酶发现以来的这50年,是真核生物转录的生物化学研究和真核转录因子鉴定的黄金年代,无数研究人员探索解决了真核生物转录过程中的一个又一个问题,但是科学的迷人之处就在于,答案越多,问题也会越多。Pol II延伸因子的发现史也带来了很多新问题,例如,转录因子如Elongin和ELL如何与转录的Pol II相互作用以抑制其停顿的?NELF和DSIF是如何诱导停顿的?Pol II是如何通过P-TEFB和SEC的作用从停顿中释放出来的?延伸因子如SII、Elongin、SEC和LEC的生物学作用是什么?这些因子在多大程度上起着一个普遍的延伸调节作用而不是基因特异性延伸调节作用?等等。或许再过50年,这些问题都会解决,但是现在,我们仍需前赴后继,寻找答案。
原文链接:
https://doi.org/10.1038/s41594-019-0283-1
制版人:珂
参考文献
1. VANNINI A, CRAMER P. Conservation between the RNA polymerase I, II, and III transcription initiation machineries[J].Molecular Cell, 2012, 45 (4) :439-46.
2. 严意华,林颖韬,胡雪峰. 真核生物RNA聚合酶的结构和功能概述[J]. 生物学教学, 2018,43(11),2-4.
来源:bioartplants BioArt植物
原文链接:http://mp.weixin.qq.com/s?__biz=MzU3ODY3MDM0NA==&mid=2247493442&idx=2&sn=1e3b868e260de113344fbf0a05f4f7ba&chksm=fd737d25ca04f43321f86d2a9f5a33ffce1d2c19c3f67ff14de37a8122b4c730063efb02dd09&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn
RNA编辑

微RNA影响家畜驯化

基于细胞外RNA的疾病诊断技术 | “自然展望-细胞外RNA”增刊
速览:阳光杀菌确实有效;外星生命可能是紫色的




新型基因芯片,可快速准确分析RNA表达模式
脊椎动物RNA病毒研究取得重要突破

抗体还能沉淀出RNA,没想到吧!(蛋白-RNA)--RIP实验解析

人类大脑纺锤形神经元转录图谱获解析

RNA编辑疗法加速发展
丙型肝炎RNA


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号