科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2020-07-22
来源:中国生物物理学会
另外,2020年6月15日,颜宁及杨洪远共同通讯在Cell 在线发表题为“Structural Basis of Low-pH-Dependent Lysosomal Cholesterol Egress by NPC1 and NPC2”的研究论文,该研究揭示了低pH依赖性胆固醇从NPC2传递到NPC1跨膜(TM)域的分子基础。在pH 8.0时,在纳米光盘和去污剂中分别获得3.6Å和3.0Å分辨率的NPC1类似结构,揭示了连接N端结构域(NTD)和跨膜固醇传感结构域(SSD)的隧道结构;在pH 5.5时,NTD表现出两个构象,表明胆固醇向隧道输送的运动。在通道的膜边界发现了一个假定的胆固醇分子,TM2向SSD上的表面袋形成。最后,在pH 5.5时获得了分辨率为4.0Å的NPC1-NPC2复合物的结构,阐明了胆固醇从NPC2转移到NPC1(NTD)的分子基础。
2020年6月8日,颜宁团队在PNAS 在线发表题为“Employing NaChBac for cryo-EM analysis of toxin action on voltage-gated Na+ channels in nanodisc”的研究论文,该研究介绍在洗涤剂胶束和纳米圆盘中NaChBac的单粒子冷冻电子显微镜(cryo-EM)分析。在两种条件下,NaChBac的构象与潜在灭活的NavAb的构象几乎相同。确定纳米光盘中NaChBac的结构使研究人员能够检查脂质双层中Nav通道的门控修饰剂毒素(GMT)。为了研究哺乳动物Nav通道中的GMT,该研究生成了一个嵌合体,其中Nav1.7的第二个电压感测域中S3和S4区段的细胞外片段替换了NaChBac中的相应序列。此解决方案可实现毒素对接的可视化。因此,NaChBac可以用作膜环境中GMT与Nav通道之间相互作用的结构研究的便捷替代品。
2020年5月13日,颜宁等团队在Nature 在线发表题为”Structural basis for catalysis and substrate specificity of human ACAT1“的研究论文,该研究介绍了人类ACAT1的冷冻电子显微镜结构。每个protomer都由九个跨膜段组成,这些段包围了一个胞质通道和一个在预计的催化位点会聚的跨膜通道。结构指导的突变分析的证据表明,酰基辅酶A通过细胞质通道进入活性位点,而胆固醇可能从侧面通过跨膜通道进入。这种结构和生化特征有助于合理化ACAT1对不饱和酰基链的偏好,并提供对MBOAT家族中酶催化机制的见解(点击阅读)。2020年5月13日,颜宁等团队在Nature 在线发表题为”Structure and mechanism of human diacylglycerol O-acyltransferase 1“的研究论文,该研究介绍了人类DGAT1的冷冻电子显微镜结构。每个DGAT1都有9个跨膜螺旋,其中8个形成保守的结构折叠,将其命名为MBOAT折叠。DGAT1中的MBOAT折叠在膜中形成一个中空腔室,该腔室包围着高度保守的催化残留物。该腔室有两个底物,脂肪酰基辅酶A和二酰基甘油的单独入口。 DGAT1可以同型二聚体或同型四聚体形式存在,两种形式具有相似的酶活性。DGAT1的N末端与邻近的protomer相互作用,而这些相互作用是酶促活性所必需的(点击阅读)。
本文转载自公众号“iNature”(Plant_ihuman)
中国生物物理学会官方订阅号,为BSC会员及生物物理领域专业人士服务。
投稿及授权请联系:bscoffice@bsc.org.cn。
微信号:BSC-1979

来源:BSC-1979 中国生物物理学会
原文链接:http://mp.weixin.qq.com/s?__biz=MzA4NjE1NTQ5Mg==&mid=2649501635&idx=1&sn=276de5400f51243e3ab14303fd5be894&chksm=87d58648b0a20f5ed07822522d4820771bcbabdfb8442ff70ee8aac3d2acd50fd3c84c63397b&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn
俄研究用新方式改善基因疗法效果
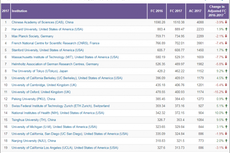
中科院位列2018自然指数榜单全球十大科研机构首位
【科技前沿】Cell:颜宁/尹航提出“饿死疟原虫”新思路
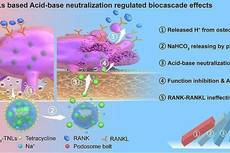
“小苏打”脂质体治疗骨质疏松

颜宁团队连续发表4篇Nature/Cell等成果,系统介绍钙离子通道蛋白调控机制

颜宁团队利用单粒子冷冻显微镜成功揭示:嵌入到脂质体中的膜蛋白的精细化结构

新突破! 二维实验系统成功应用于四维材料研究
【共同关注】女科学家去哪儿了?



复旦大学最新Science,把仿生纳米脂质体推向更广谱

颜宁团队连续发表5篇Cell/Nature,提出了通用的解析膜蛋白的方法


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号