科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2019-08-16
来源:ScienceAAAS
中科院青促会特邀述评人 晁青(中科院植物研究所)
评述论文:Teosinte ligule allele narrows plant architecture and enhances high-density maize yields(Science 16 Aug 2019: Vol 365, Issue 6454)
玉米是世界第一大粮食作物,全世界玉米总产量已经超过10亿吨,占世界粮食总产量的41%,对全球的粮食安全起着举足轻重的作用。在过去的半个世纪,玉米的产量因为种植密度的增加而得到提高[1]。但是,种植密度的增加需要更加紧凑的株型,主要是更小的叶夹角产生向上的叶片[2,3]。叶夹角的大小直接影响玉米群体冠层的光能利用率和群体的光合效率,也决定了种植密度的高低。
玉米的叶夹角主要由分隔叶片(Blade)和叶鞘(Sheath)的叶舌(Ligule)和叶耳(Auricle)(图1)决定。已有研究鉴定叶夹角主要由以下三类基因的突变决定:第一类是无叶舌和叶耳的突变,如liguleless1(lg1),lg2和ligulelessnarrow[4];第二类是调节叶耳的大小的突变,如油菜素内酯(BR)激素信号途径[5,6];第三类是主脉的厚度的变化,如缺乏主脉的drooping leaf(drl)[7]。尽管这些突变可以改变叶夹角,但由于其多效性(如松垂的叶子),不能直接用于提高玉米的产量。
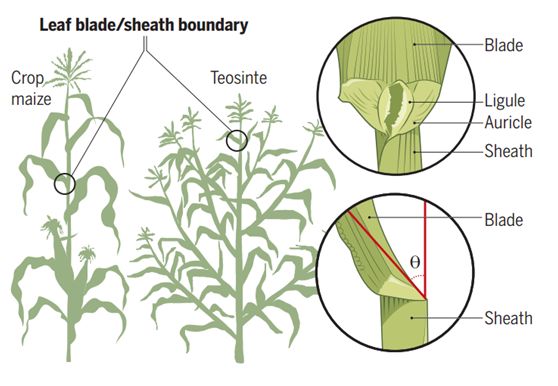
图1. 玉米(Crop maize)和大刍草(Teosinte)的叶夹角组成机制
本期的《科学》杂志在线发表了中国农业大学田丰教授团队在玉米叶夹角方面的研究。科研人员利用玉米野生种——大刍草(CIMMT 8759)与玉米自交系(W22)为亲本杂交衍生得到的深入系群体,对叶夹角进行数量性状位点(QTL)的定位,并对玉米的两个主效QTLUPA1(Upright Plant Architecture1)和UPA2进行了精细定位。
结果发现,UPA2造成叶夹角的差异主要是由位于240bp非编码区域内的2个核苷酸的插入/缺失造成的,而这一插入只存在于野生种大刍草中,在人工驯化的自交系中已经完全缺失。它可以作为顺式调控元件调控位于下游9.5 Kb的基因ZmRAV1。而上文提到的dr1l编码的蛋白DRL1蛋白可以更好地结合在大刍草的等位基因上,并且与玉米控制叶舌的LG1互作,从而抑制LG1对ZmRAV1的激活。另一个数量性状位点UPA1的功能基因是BR途径的合成基因brd1。而ZmRAV1可以调控brd1的表达,最终导致了大刍草中brd1的下调。进而降低叶环处内源BR水平,影响叶耳细胞的增殖,最终导致叶夹角减小,株型趋于紧凑(图2)。
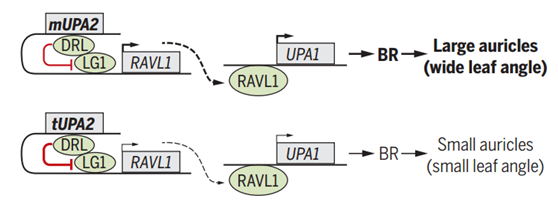
图2. 玉米叶夹角形成的调控机制
玉米是大约9000年前由分布于墨西哥西南部的大刍草驯化而来。在其驯化过程中,由于遗传瓶颈效应和选择作用,玉米丢失了大刍草~30%的遗传多样性,其丢失的遗传多样性中可能包含可用于现代育种的优良等位基因。
田间试验表明,UPA2的大刍草近等基因系在密植条件下具有显著的增产效应。借助分子标记辅助选择,该研究将UPA2的大刍草等位基因回交导入到了优良玉米杂交种农大108双亲中,获得了携带UPA2大刍草等位基因的改良农大108。
田间密植试验显示,改良的农大108在密植条件下玉米籽粒产量显著增加(图3)。这些结果都说明:UPA2野生等位变异在当前密植高产育种中可能具有重要的利用价值,充分利用玉米野生种质资源是解决目前育种遗传基础狭窄的重要途径。
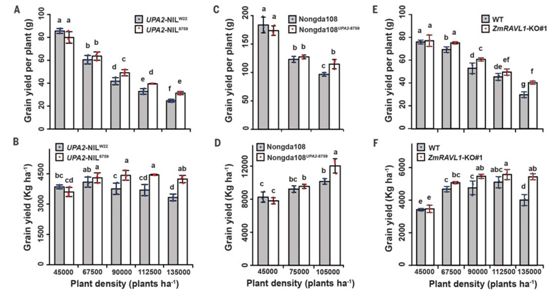
图3. 大刍草的UPA2和ZmRAVL1的编辑可以提高密植条件下的玉米产量
该研究从玉米野生祖先种大刍草中克隆了控制玉米紧凑株型、密植增产的关键基因,建立了玉米紧凑株型的分子调控网络,为玉米理想株型分子育种、培育密植高产品种提供了理论和实践基础。
述评人简介:晁青博士,中国科学院植物研究所副研究员。主要研究方向为玉米的光合碳代谢、植物的维管发育和抗病等方面的研究。参考文献:
1. Duvick D. Genetic progress in yield of United States maize (Zea mays L.). Maydica, 2005, 50: 193-202.
2. Pendleton J, Smith G, Winter S, et al. Field investigations of the relationships of leaf angle in corn (Zea mays L.) to grain yield and apparent photosynthesis. Agronomy Journal, 1968, 60: 422-424.
3. Lambert R, Johnson R. Leaf Angle, Tassel morphology, and the performance of maize Hybrids. Crop Science, 1978, 18: 499-502.
4. Richardson A, Hake S. Drawing a line: Grasses and boundaries. Plants, 2019, 8: 4.
5. Tong H, Liu L, Jin Y, et al. DWARF AND LOW-TILLERING acts as a direct downstream target of a GSK3/SHAGGY-like kinase to mediate brassinosteroid responses inrice. The Plant Cell, 2012, 24: 2562-2577.6. Hong Z, Ueguchi-Tanaka M, Matsuoka M. Brassinosteroids and rice architecture. Journal of Pesticide Science, 2004, 29: 184-188.
7. Strable J, Wallace JG, Unger-Wallace E, et al. Maize YABBY genes drooping leaf1 and drooping leaf2 regulate plant architecture. The Plant Cell, 2017, 29: 1622-1641.
来源:Science-AAAS ScienceAAAS
原文链接:http://mp.weixin.qq.com/s?__biz=MzI3NDY3NzQ2Mg==&mid=2247487804&idx=2&sn=43fae63c0af1a1c7886b93746d6407e9&chksm=eb113a85dc66b3935eccd601ee726aff28a26a1cce46eaef29cfe23894e7ea6e83c374d534e7&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn
四师农科所三区科特派团队到67团进行科技服务


心理所发现伏隔核激活与愉快体验共享遗传信息

冰岛人民的遗传信息让你认识不一样的尼安德特人

打破时间限制,科学家在170万年前的犀牛牙齿中提取出遗传信息

国际最新研究:斑鬣狗DNA遗传信息反映社会地位等级

研究发现,遗传影响儿童观察模式和心智发展

每日科普 | 如果遗传信息被泄漏,后果将远超您的想象
玉米薄饼

古DNA技术:挖掘古代人类遗传信息的利器
科学家最新发现,RNA在DNA“失能”时主持生命密码传递工作


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号