科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2019-09-09
来源:青塔
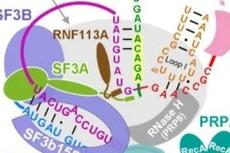
剪接体通过两个顺序的酯交换反应,分支和外显子连接来执行真核前体信使RNA(pre-mRNA)剪接以去除非编码内含子。该过程的保真度基于对内含子中的保守序列的识别以及该多兆达尔顿机器的动态组成和结构重排。自2015年内源性裂殖酵母剪接活性剪接活性位点的原子可视化以来,其他剪接体中间体的高分辨率冷冻电镜结构开始揭示其分子机制。剪接体的结构生物学的最新进展使其更清楚其组装,活化,解体和外显子连接的机制。这些离散的结构图像一起产生剪接体的分子编排。
近日,西湖大学施一公团队(一作清华万蕊雪)在Current Opinion in Structural Biology上发表了题为Molecular choreography of pre-mRNA splicing by the spliceosome的综述性文章。该文章着重于酵母剪接体结构生物学的最新进展。


由于长度限制,酵母和人类剪接体之间的结构比较不包括在本综述中。但两个剪接体的关键特征是相同的。

前mRNA剪接在1977年有明确的记录。小核糖核蛋白(SnRNPs)在剪接中的潜在作用立即被观察到。负责剪接的分子机制,称为剪接小体,于1985年分离。剪接体包括五个不同的富含尿苷的RNPs(U1、U2、U4、U5和U6 snRNPs)、19个复合物(NTC)和NTC相关复合物(NTR)的约18个蛋白质、大约16个剪接因子、8个依赖RNA的ATP酶/螺旋酶,以及许多调节和辅助蛋白。
pre-mRNA 的每个内含子被5'-外显子和3'-外显子包围,并含有至少3个保守的RNA元件:5'-剪接位点(5'SS),分支点序列(BPS)和3'。-splice site(3'SS)。剪接进行两步:分支和外显子连接。支化反应产生游离的5'-外显子和内含子套索-3'-外显子中间体; 外显子连接加入5'-外显子和3'-外显子并释放内含子套索。两种反应都被称为M1和M2的两种二价金属离子催化,它们通过来自剪接体活性位点的U6 snRNA的保守核苷酸进行协调。

△剪切过程
完整的剪接周期由剪接体的四个连续阶段组成:组装,激活,催化和拆卸。组装开始于U1 snRNP识别5'SS,SF1识别BPS,U2AF识别3'SS。U2 snRNP然后替换SF1以生成A复合物,其募集U4 / U6.U5 tri-snRNP以形成完全组装的前B复合物。在剪接体激活期间,前B复合物首先脱落U1 snRNP成为B复合物。在Brr2作用后,U4 snRNP解离,募集NTC和NTR复合物以形成活化的剪接体(Bact)。
为了发生剪接,Bact复合物被ATP酶/解旋酶Prp2进一步活化,成为B *复合物,其中支化反应在步骤I因子Cwc25和Yju2存在下进行。得到的C复合物需要Prp16驱动的重塑成为C *复合物,其中外显子连接在步骤II因子Prp18和Slu7的存在下发生,产生后催化剪接体(P复合物)。剪接体的解体分两步进行:通过Prp22介导的连接外显子的释放形成内含子套索剪接体(ILS)和通过Prp43分解ILS复合物。本综述着重于酵母剪接体结构生物学的最新进展。由于长度限制,酵母和人类剪接体之间的结构比较不包括在本综述中。但两个剪接体的关键特征是相同的。
原文链接:
https://www.sciencedirect.com/science/article/abs/pii/S0959440X19300430#!
来源:cingta-com 青塔
原文链接:http://mp.weixin.qq.com/s?__biz=MzI5MjIwMzk1MQ==&mid=2247507517&idx=4&sn=139b73554710d9f6d58c6002612ee23d&chksm=ec0662fddb71ebeb04473f57d760e70b7e2d10604394f374aa6a429d57749aa2f5bcd1a9d23c&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn

施一公:生物科学男神教授

西湖大学在次要剪接体领域再获突破
西湖大学施一公研究组Science:首个人源次要剪接体的电镜结构

在梦想与现实不可调和时,施一公为什么选择科研之路

施一公为西湖大学请辞清华副校长
【大师讲堂】外显子拼接是如何完成的?

次要剪接体:不次要的生命“剪辑师”

【科技前沿】施一公等揭示剪接体核酶活性中心有序折叠的机制和后续重塑的动力机制

西湖大学明年成立,5年后科研水平比肩清华

施一公再刊新作(一作万蕊雪),阐述酵母剪接体结构生物学最新进展


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号