

科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2020-11-17
群落 (Biocoenosis)或称为“生物群落”。指的是生态学中,在一个群落生境内相互之间具有直接或间接关系的所有生物,或曰生物的总合。也有人作如下定义:生存在一起并与一定的生存条件相适应的动植物的总体。群落生境是群落生物生活的空间。一个生态系统则是群落和群落生境的系统性相互作用。一个群落的生物物种占据不同的小生境。一个群落的生物相互之间有着不同的关系。
概念来源1877年,德国动物学家Karl Möbius创造了biocenosis一词(Möbius 1877),描述生活在一个生境(habitat)中相互作用的有机体,认为每一个生物群落都“支持着一定数量的有生命的生物”,强调在合适的条件下可能产生过量的后代,但由于空间和食物是有限的,“群落中个体总数不久又会回到它以前的适中状态”,这使人回想起马尔萨斯关于人口与它们受资源限制的理论。但在英语中biocenosis一词很少使用,而常用Ecological community,指占据同一地理区域的二个或以上不同种类的物种的集合。
生物群落作为一般名词时与群落同义。生物群落一词,只限于强调沿袭克列门茨和谢尔福德(F.E.Clements & V.E.Shelford,1939)的考虑方法。克列门茨认为,植物群落并不是个体和种的组合,而是用生长型(growth form)为代表的生态群组合,这种必须以生物个体为准据的新阶段的有机体,称之为复合生物(complex organism)。谢尔福德(1912)对动物曾使用了生理活动型这个名词,它相当于植物的生长型,并称之为生态种群(mores,用复数形,单数形为mos,涵义不同),它的组合即是动物群落。以后把植物和动物作为mune的机能生活形的集群,而进一步强调了统一性。演替是复合生物的发育、成长,相当于它的成体的顶极群落是仅仅由气候决定的,也有把这种观点特称之为生物生态学(bio-ecology)的。1
构成生物群落结构与生态系统的概念不同。后者不仅包括生物群落还包括群落所处的非生物环境,把二者作为一个由物质、能量和信息联系起来的整体。因此生物群落只相当于生态系统中的生物部分。
生物群落中的各种生物之间的关系主要有3类:
①营养关系,当一个种以另一个种,不论是活的还是它的死亡残体,或它们生命活动的产物为食时,就产生了这种关系。又分直接的营养关系和间接的营养关系。采集花蜜的蜜蜂,吃动物粪便的粪虫,这些动物与作为它们食物的生物种的关系是直接的营养关系;当两个种为了同样的食物而发生竞争时,它们之间就产生了间接的营养关系。因为这时一个种的活动会影响另一个种的取食。
②成境关系,一个种的生命活动使另一个种的居住条件发生改变。植物在这方面起的作用特别大。林冠下的灌木、草类和地被以及所有动物栖居者都处于较均一的温度、较高的空气湿度和较微弱的光照等条件下。植物还以各种不同性质的分泌物(气体的和液体的)影响周围的其他生物。一个种还可以为另一个种提供住所,例如,动物的体内寄生或巢穴共栖现象,树木干枝上的附生植物等。
③助布关系,指一个种参与另一个种的分布,在这方面动物起主要作用。它们可以携带植物的种子、孢子、花粉,帮助植物散布。
营养关系和成境关系在生物群落中具有最大的意义,是生物群落存在的基础。正是这两种相互关系把不同种的生物聚集在一起,把它们结合成不同规模的相对稳定的群落。
基本特征1.具有一定的动态特征
2.具有一定的边界特征
3.具有一定的分布范围
4.具有一定的结构
5.具有一定的种类组成
6.具有一定的群落环境
7.不同物种之间的相互影响
8.各物种不具有同等的群落学重要性
基本原则⒈一个群落环境的小环境越是多样,那么其中的群落就越是物种丰富。(热带雨林)
⒉一个群落环境的非生物因素和全球平均值相差越大,其物种也会越少,但个体数目却会越多。这个区域的群落通常是高特异性的。(盐湖,深海)
⒊一个群落环境的生存环境越是缓慢连续的交替改变,群落也会越是丰富。(珊瑚礁)
在生物群落中,各个种群占据了不同的空间,使群落具有一定的结构。群落的结构包括垂直结构和水平结构。
相对稳定了的生物群落其重要特征之一,是具有一定的空间结构。群落中各种生物在空间上的配置状况,即为群落的结构。群落结构包括形态方面的结构和生态方面的结构,前者包括垂直结构和水平结构,后者指层片结构。
结构分类包括空间结构、时间组配和种类结构。
空间结构不同生活型的植物(乔木、灌木、草本)生活在一起,它们的营养器官配置在不同高度(或水中不同深度),因而形成分层现象。分层使单位面积上可容纳的生物数目加大,使它们能更完全、更多方面地利用环境条件,大大减弱它们之间竞争的强度;而且多层群落比单层群落有较大的生产力。
分层现象在温带森林中表现最为明显,例如温带落叶阔叶林可清晰地分为乔木、灌木、草本和苔藓地衣(地被)4层。热带森林的层次结构最为复杂,可能有的层次最为发育,特别是乔木层,各种高度的巨树、一般树和小树密集在一起,但灌木层和草本层常常不很发育。草本群落一样地分层,尽管层次少些(通常只分为草本层和地被层)。
群落不仅地上分层,地下根系的分布也是分层的。群落地下分层和地上分层一般是相应的;乔木根系伸入土壤的最深层,灌木根系分布较浅,草本植物根系则多集中土壤的表层,藓类的假根则直接分布在地表。
生物群落结构的垂直分层与光照条件密切相关,每一层的植物适应于该层的光照水平,并降低下层的光强度。在森林中光强度向下递减的现象最为明显。最上层树处于全光照之中,平均说来,到达下层小树的光只有上层树(全光照)的10~50%,灌木层只有5~10%,而草本层则只剩1~5%了。随着光照强度的变化,温度、空气湿度也发生变化。
每一层植物和被它们所制约的小气候为生活于其中的特有动物创造一定的环境,因此动物在种类上也表现出分层现象,不同的种类出现于不同层次,甚至同一种的雌雄个体,也分布于不同的层次。例如,在森林中可以区分出3组鸟种:在树冠中采食的,接近地面的,以及生活在其间的灌木和矮树簇叶中的。
林地也由于枯枝落叶层的积累和植物对土壤的改造作用,创造了特殊的动物栖居环境。较高的层(草群,下木)为吃植物的昆虫、鸟类、哺乳动物和其他动物所占据。在枯枝落叶层中,在腐烂分解的植物残体、藓类、地衣和真菌中,生活着昆虫、蜱、蜘蛛和大量的微生物。在土壤上层,挤满了植物的根,这里居住着细菌、真菌、昆虫、蜱、蠕虫。有时在土壤的某种深度还有穴居的动物。
当然,也存在一些层外生物,它们不固定于某一个层。例如藤本植物、附生植物,以及从一个层到另一个层自由活动的动物。它们使划分层次困难化;在结构极其复杂的热带雨林中经常见到这种情况。
因为下层生物是在上层植物遮荫所形成的环境中发育起来的,所以生物群落中不同层的物种间有密切的相互作用和相互依赖关系。群落上层植物强烈繁生,相应地下层植物的密度就会降低;而如果由于某种原因上层植物变得稀疏,下层的光照、热等状况得到改善,同时土壤中矿物养分因释放加强而增高,下层植物发育便会加强。下层的繁茂生长也对动物栖居者有利。这种情况特别反映在森林群落中,哪里乔木层稀疏便会导致那里的灌木或喜光草本植被的丰富繁生。而乔木层的完全郁闭,有时甚至抑制最耐荫的草本和藓类。
生物群落不仅有垂直方向的结构分化,而且还有水平方向的结构分化。群落在水平方向的不均匀性表现为以斑块出现;在不同的斑块上,植物种类、它们的数量比例、郁闭度、生产力以及其他性质都有不同。例如在一个草原地段,密丛草针茅是最占优势的种类,但它并不构成连续的植被,而是彼此相隔一定的距离(30~40厘米)分布的。各个针茅草丛之间的空间,则由各种不同的较小的禾本科植物和双子叶杂类草占据着,并混有鳞茎植物。但其中的某些植物也出现于针茅草丛的内部。因此,伴生少数其他植物的针茅草丛同针茅草丛之间生长有其他草类的空隙,它们在外貌、在种间数量关系和质量关系上都有很明显的不同。但它们的差别与整个植物群落(针茅草原)比较起来,是次一级的差别,而且是不很明显的和不稳定的。在森林中,在较阴暗的地点和较明亮的地点,也可以观察到在植物种类的组成和数量比例方面以及其他方面的类似差异。群落内水平方向上的这种不一致性,叫做群落的镶嵌性。这种不一致性在某些情况下是由群落内环境的差别引起的,如影响植物种分布的光强度不同或地表有小起伏;在某些情况下是由于共同亲本的地下茎散布形成的植物集群所引起;在另外的情况下,它们可能由种之间的相互作用引起,例如在寄主种的根出现的地方形成斑块状的寄生植物。动物的活动有时也是引起不均一性的原因。植物体通常不是随机地散布于群落的水平空间,它们表现出成丛或成簇分布。许多动物种群,不论在陆地群落或水生群落,也具有成簇分布的性质。相比之下,有规则的分布是比较不常见的。某些荒漠中灌木的分布、鸣禽和少数其他动物的均匀分布是这种有规则分布的例子。
1.垂直结构
群落的垂直结构指群落在垂直方面的配置状态,其最显著的特征是成层现象,即在垂直方向分成许多层次的现象。
群落的成层性包括地上成层和地下成层。层的分化主要决定于植物的生活型,生活型不同,植物在空中占居的高度以及在土壤中到达的深度就不同,水生群落则在水面以下不同深度形成物种的分层排列,这样就出现了群落中植物按高度(或深度)配置的成层现象。
成层现象在森林群落表现最为明显,而以温带阔叶林和针叶林的分层最为典型,热带森林的成层结构则最为复杂。一般按生长型把森林群落从顶部到底部划分为乔木层、灌木层、草本层和地被层(苔藓地衣)四个基本层次,在各层中又按植株的高度划分亚层,例如热带雨林的乔木层通常分为三个亚层。草本群落则通常只有草本层和地被层。
在层次划分时,将乔木和其他生活型植物不同高度的幼苗划入实际所逗留的层中,生活在各层中的地衣、藻类、藤本等层间植物通常也归入相应的层中。
群落的地下分层和地上分层一般是相应的。森林群落中的乔木根系为分布到土壤的深层,灌木根系较浅,草本植物的根系则大多分布在土壤的表层。草本群落的地下分层比地上分层更为复杂。
群落的成层性保证了植物在单位空间中更充分利用自然环境条件。如在发育成熟的森林中,上层乔木可以充分利用阳光,而林冠下为那些能有效利用弱光的下木所占居,林下灌木层和草本层能够利用更微弱的光线、草本层往下还有更耐荫的苔藓层。
生物群落中动物的分层现象也很普遍。动物分层主要与食物有关,其次还与不同层次的微气候条件有关。如东欧亚大陆北方针叶林区,在地被层和草本层中,栖息着两栖类、爬行类、鸟类(丘鹬、楱鸡)、兽类(黄鼠)和各种鼠形啮齿类;在森林的下层——灌木林和幼林中,栖息着莺、苇莺和花鼠等;在森林的中层栖息着山雀、啄木鸟、松鼠和貂等,而在树冠层则栖息着柳莺、交嘴和戴菊等。
水域中某些水生动物也有分层现象,这主要决定于阳光、温度、食物和含氧量等。比如湖泊,在一年当中湖水没有循环流动的时候,浮游动物都表现出明显的垂直分层现象,它们多分布在较深的水层,在夜间则上升到表层来活动,这是因为浮游动物一般都是趋向弱光的。
2.水平结构
群落的水平结构指群落的水平配置状况或水平格局,其主要表现特征是镶嵌性。
镶嵌性即植物种类在水平方向不均匀配置,使群落在外形上表现为斑块相间的现象。具有这种特征的群落叫做镶嵌群落。在镶嵌群落中,每一个斑块就是一个小群落,小群落具有一定的种类成份和生活型组成,它们是整个群落的一小部分。例如,在森林中,林下阴暗的地点有一些植物种类形成小型的组合,而在林下较明亮的地点是另外一些植物种类形成的组合。这些小型的植物组合就是小群落。内蒙古草原上锦鸡儿灌丛化草原是镶嵌群落的典型例子。在这些群落中往往形成1~5米左右的锦鸡儿丛,呈园形或半园形的丘阜。这些锦鸡儿小群落内部由于聚集细土、枯枝落叶和雪,因为具有良好的水分和养分条件,形成一个局部优越的小环境。小群落内部的植物较周围环境中返青早,生长发育好,有时还可以遇到一些越带分布的植物。
群落镶嵌性形成的原因,主要是群落内部环境因子的不均匀性,例如小地形和微地形的变化,土壤温度和盐渍化程度的差异,光照的强弱以及人与动物的影响。在群落范围内,由于存在不大的低地和高地因而发生环境的改变形成镶嵌,这是环境因子的不均匀性引起镶嵌性的例子。由于土中动物,例如田鼠活动的结果,在田鼠穴附近经常形成不同于周围植被的斑块,这是动物影响镶嵌性的例子。
3.层片结构
层片一词系瑞典植物学家加姆斯发明创造。他起初赋于这一概念以三个方面的内容,即把层片划分为三级:一级层片,即同种个体的组合;二级层片,即同一生活型的不同植物的组合;三级层片,即不同生活型的不同种类植物的组合。一般群落学研究中使用的层片概念,均相当于加姆斯的二级层片,即每一个层片都是由同一生活型的植物所组成。
生活型是植物对外界环境适应的外部表现形式,同一生活型的植物不但体态上是相似的,而且在形态结构、形成条件、甚至某些生理过程也具相似性。如今广泛采用的生活型划分是郎基耶尔的系统。他按照休眠芽在不良季节的着生位置把植物的生活型分成五大类群,高位芽植物(25厘米以上),地上芽植物(25厘米以下)、地面芽植物(位于近地面土层内)、隐芽植物(位于较深土层或水中)和一年生植物(以种子越冬),在各类群之下再细分为30个较小的类群。我国植被学著作中采用的是按体态划分的生活型系统,该系统把植物分成木本植物、半木本植物、草本植物、叶状体植物四大类别,再进一步划分成更小的或低级的单位。对于层片的划分,可以根据研究的需要,分别使用上述系统中的高级划分单位或低级单位。
层片作为群落的结构单元,是在群落产生和发展过程中逐步形成的。它的特点是具有一定的种类组成,它所包含的种具有一定的生态生物学一致性,并且具有一定的小环境,这种小环境是构成植物群落环境的一部分。
需要说明一下层片与层的关系问题。在概念上层片的划分强调了群落的生态学方面,而层次的划分,着重于群落的形态。层片有时和层是一致的,有时则不一致。例如分布在大兴安岭的兴安落叶松纯林,兴安落叶松组成乔木层,它同时也是该群落的落叶针叶乔木层片。在混交林中,乔木层是一个层,但它由阔叶树种层片和针叶树种层片两个层片构成。在实践中,层片的划分比层的划分更为重要,但划分层次往往是区分和分析层片的第一步。
和层结构一样,群落层片结构的复杂性,保证了植物全面利用生境资源的可能性,并且能最大程度地影响环境,对环境进行生物学改造。2
时间组配组成群落的生物种在时间上也常表现出“分化”,即在时间上相互“补充”,如在温带具有不同温度和水分需要的种组合在一起:一部分生长于较冷季节(春秋),一部分出现于炎热季节(夏)。例如,在落叶阔叶林中,一些草本植物在春季树木出叶之前就开花,另一些则在晚春、夏季或秋季开花。随着不同植物出叶和开花期的交替,相联系的昆虫种也依次更替着:一些在早春出现,另一些在夏季出现。鸟类对季节的不同反应,表现为候鸟的季节性迁徙。生物也表现出与每日时间相关的行为节律:一些动物白天活动;另一些黄昏时活动;还有一些在夜间活动,白天则隐藏在某种隐蔽所中。大多数植物种的花在白天开放,与传粉昆虫的活动相符合;少数植物在夜间开花,由夜间动物授粉。许多浮游动物在夜间移向水面,而在白天则沉至深处远离强光,但是不同的种具有不同的垂直移动模式和范围、潮汐的复杂节律控制着许多海岸生物的活动。土壤栖居者也有昼夜垂直移动的种类。
种类结构每一个具体的生物群落以一定的种类组成为其特征。但是不同生物群落种类的数目差别很大。例如,在热带森林的生物群落中,植物种以万计,无脊椎动物种以10万计,脊椎动物种以千计,其中的各个种群间存在非常复杂的联系。
生物群落
冻原和荒漠群落的种数要少得多。根据苏联学者Б.А.季霍米罗夫的资料,在西伯利亚北部的泰梅尔半岛的冻原生物群落中共有139种高等植物,670种低等植物,大约1000种动物和2500种微生物。与此相应。这些生物群落的生物量和生产力,也比热带森林小得多。
生物群落中生物的复杂程度用物种多样性这一概念表示。多样性与出现于某一地区的生物种的数量有关,也与个体在种之间的分布的均匀性有关。例如,两个群落都含有5个种和100个个体,在一个群落中这100个个体平均地分配在全部5个种之中,即每1个种有20个个体,而在另一个群落中80个个体属于1个种,其余20个个体则分配给另外的4个种,在这种情况下,前一群落比后一群落的多样性大。
在温带和极地地区,只有少数物种很常见,而其余大多数物种的个体很稀少,它们的种类多样性就很低;在热带,个体比较均匀地分布在所有种之间,相邻两棵树很少是属于同种的(热带雨林),种类多样性就相对较高。群落的种类多样性决定于进化时间、环境的稳定性以及生态条件的有利性。热带最古老,形成以来环境最稳定,高温多雨气候对生物的生长最为有利,以生物群落的种类多样性最大。在严酷的冻原环境中,情况相反,所以种类多样性低。
每种植物在群落中所起的作用是不一样的。常常一些种以大量的个体,即大的种群出现;而另一些种以少量的个体,即小的种群出现。个体多而且体积较大(生物量大)的植物种决定了群落的外貌。例如,绝大多数森林和草原生物群落的一般外貌决定于一个或若干个植物种,如中国山东半岛的大多数栎林决定于麻栎,燕山南麓的松林决定于油松,内蒙古高原中东部锡盟的针茅草原决定于大针茅或克氏针茅等。在由数十种甚至百余种植物组成的森林中,常常只有一种或两种乔木提供90%的木材。群落中的这些个体数量和生物量很大的种叫做优势种,它们在生物群落中占居优势地位。优势种常常不止一个,优势种中的最优势者叫建群种,通常陆地生物群落根据建群植物种命名,例如,落叶阔叶林、针茅草原、泥炭藓沼泽等。建群种是群落的创建者,是为群落中其他种的生活创造条件的种。例如,云杉在泰加带形成稠密的暗针叶林,在它的林冠下,只有适应于强烈遮荫条件,高的空气湿度和酸性灰化土条件的植物能够生活;相应于这些因素,在云杉林中还形成特有的动物栖居者。因此在该情况下云杉起着强有力的建群种的作用。
松林中的建群种是松树,但与云杉相比,它是较弱的建群种,因为松林树干稀疏,树冠比较不密接,比较透光,它的植物和动物的种类组成远比云杉林丰富和多样。在松林中甚至见到能生活在林外环境中的植物。
温带和寒带地区的生物群落中,建群种比较明显;无论森林群落、灌木群落、草本群落或藓类群落,都可以确定出建群种(有时不止一个)。亚热带和热带,特别是热带的生物群落,优势种不明显,很难确定出建群种来。除优势种外,个体数量和生物量虽不占优势但仍分布广泛的种是常见种;个体数量极少,只偶尔出现的种是偶见种。
生物群落中的大多数生物种,在某种程度上与优势种和建群种相联系,它们在生物群落内部共同形成一个物种的综合体,叫做同生群。同生群也是生物群落中的结构单位。例如一个优势种植物,和与它相联系的附生、寄生、共生的生物以及以它为食的昆虫和哺乳动物等共同组成一个同生群。
生活在一个群落中的多种多样的生物种,是在长期进化过程中被选择出来能够在该环境中共同生存的种。它们中每一个占据着独特的小生境,并且在改造环境条件、利用环境资源方面起着独特的作用。群落中每一个生物种所占据的特定的生境和它执行的独特的功能的结合,叫做生态位。因此,一个生物群落的物种多样性越高,其中生态位分化的程度也越高。
影响要素生物因素群落结构总体上是对环境条件的生态适应,但在其形成过程中,生物因素起着重要作用,其中作用最大的是竞争与捕食。
竞争对群落结构的影响
竞争对群落结构的形成有重要影响。对种间竞争在形成群落结构的作用问题上,最直接的证据可能是在自然群落中对物种进行引进或去除实验,Schoner和 Cornell 就分别对种间竞争研究的文献进行过统计(分别达184和72 例研究),平均有90%的例证说明有种间竞争,表明自然群落中竞争是相当普遍的。他们的结果还表明,海洋生物中有种间竞争的比例较陆地生物多;大型生物间比小型生物间高;而植食性昆虫中竞争比例低,因为绿色植物到处都有,较丰富,很少被一食而空,所以为食物资源而竞争的可能性比较小。
已有证据表明,竞争是群落形成的一个重要驱动因素。但竞争的重要性在多个群落间显然是不同的,而且常常只是影响物种之间相互作用的一小部分。
那么为什么许多调查结果显示的竞争往往是不强烈的呢?
一般认为:① 自然选择可能已有效地通过生态位划分而避免了竞争(或者抹去了过去竞争的痕迹);② 在一个环境斑块中,具有强竞争力的物种共存,因为它们并不利用相同的资源;③ 物种也许仅仅在种群爆发、资源短缺时才发生竞争。
捕食对群落结构的影响
捕食对形成群落结构的影响,视捕食者是泛化种还是特化种而不同。对泛化种来说,捕食使种间竞争缓和,并促进多样性提高。但当取食强度过高时,物种数亦随之降低。对特化种来说,随被选食的物种是优势种还是劣势种而异。如果被选择的是优势种,则捕食能提高多样性,如果捕食者喜食的食竞争力弱的劣势种,多样性就会随着呈现型下降趋势。
干扰对群落结构的影响
干扰是自然界的普遍现象。干扰不同于灾难,不会产生巨大的破坏作用,但它经常会反复的出现,使物种没有充足的时间进化。近代多数生态学家认为干扰是一种有益的生态现象,它引起群落的非平衡特性,强调了干扰在群落结构形成和动态中的作用。
① 干扰与群落的缺口
连续的群落中出现缺口是非常普遍的现象,而缺口经常由干扰构成。森林中的缺口可能由大风、雷电、砍伐、火烧等引起;草地群落的干扰包括放牧、动物挖掘、践踏等。干扰造成群落的缺口以后,有的在没有继续干扰的条件下会逐渐地恢复,但缺口也可能被周围群落的任何一个种侵入和占有,并发展为优势者,哪一种是优胜者完全取决于随机因素,这可称为对缺口的抽彩式竞争。
但是,有些群落所形成的缺口,其物种更替是有规律性的。新打开的缺口常常被扩散能力强的一个或几个先锋种所入侵。由于它们的活动,改变了条件,促进了演替中期种的入侵,最后为顶级种所替代。在这种情况下,多样性开始较低,演替中期增加,但到顶级期往往稍有降低。
② 中度干扰假说
缺口形成的频率影响物种多样性,据此,T.W.Connell等提出了中度干扰假说,即中等程度的干扰水平能维持高多样性。其理由是:
一、在一次干扰后少数先锋物种入侵缺口,如果干扰频繁,则先锋种不能发展到演替中期,因而多样性较低;
二、如果干扰间隔期很长,使演替过程能发展到顶级期,多样性也不很高;
三、只有中等干扰程度使多样性维持最高水平,它允许更多的物种入侵和定居。
|| ||
空间异质性群落的环境不是均匀一致的,空间异质性的程度越高,意味着有更加多样的小生境,所以能允许更多的物种共存。
植物群落研究中大量资料说明,在土壤和地形变化丰富的地方,群落会有更多的物种;而平坦、同质土壤的群落多样性低。Harman 研究了淡水系统软体动物种数与空间异质性的相关性,也得出栖境质类型越多,淡水软体动物种数越多的正相关的关系结果。
MacArthur等通过研究鸟类多样性与植物物种多样性和取食多样性之间的关系,发现鸟类多样性与植物种数的相关,不如与取食——高度多样性相关明显。所以对于鸟类生活来说,植被的分层结构比物种组成更为重要。在草地和灌丛群落中,垂直结构不如森林群落明显,而水平结构,即镶嵌性和斑块性,就可能起到决定性作用。
从表上可知,每一类植物群落都是由几种生活型的植物所组成,但其中有一类生活型占优势,生活型与环境关系密切,高位芽植物占优势是温暖、潮湿气候地区群落的特征,如热带雨林群落;地面芽植物占优势的群落,反映了该地区具有较长的严寒季节,如温带针叶林、落叶林群落;地下芽植物占优势,反映了该地区环境比较湿冷,如长白山寒温带暗针叶林;一年生植物占优势则是干旱气候的荒漠和草原地区群落的特征,如东北温带草原。
|| ||
岛屿效应岛屿的物种数与面积的关系
岛屿由于与大陆隔离,生物种迁入和迁出的强度低于周围连续的大陆。许多研究证实,岛屿中的物种数目与岛屿的面积有密切关系。岛屿面积越大,岛屿上的物种数越多,岛屿面积与岛屿上物种数的关系可以用简单方程描述为:
S = cAz
或用对数表示:lgS = lgc + z (lgA)
式中:S —— 种数;
A —— 面积;
z 、c —— 两个常数。
⒉3.4.2 MacArthur 的平衡说
岛屿上的物种数虽决定于岛屿的面积,但它是物种迁入、迁出和灭亡平衡的结果,这是一种动态平衡,不断地有物种灭亡,也不断地由同种或别种的迁入而补偿灭亡的物种。
岛屿上物种的平衡关系可用图3 — 2说明,以迁入率曲线为例,当岛上无留居种时,任何迁入个体都是新的,因而迁入率高。随着留居种数加大,种的迁入率就下降。当种源库(大陆上的种)所有的种在岛上都有时,迁入率为零。灭亡率则相反,留居种数越多,灭亡率越高。迁入率取决于岛屿与大陆距离的远近和岛屿的大小,离大陆较近且面积大的岛,其迁入率高;远而小的岛,迁入率低。同样,灭亡率也受岛的大小影响。
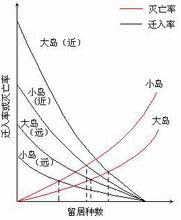 迁入率曲线与灭亡率曲线交点上的种数,即为该岛上预测的物种数。根据平衡说,可说明下列四点:
迁入率曲线与灭亡率曲线交点上的种数,即为该岛上预测的物种数。根据平衡说,可说明下列四点:
① 岛屿上的物种数不随时间而变化;
② 这是一种动态平衡,即灭亡种不断地被新迁入的种所代替;
③ 大岛比小岛能“供养”更多的种;
④ 随岛距大陆的距离由近到远,平衡点的种数逐渐降低。3
平衡学说和非平衡学说
对于形成群落结构的一般理论,有两种对立的观点,即平衡学说和非平衡学说。
平衡学说认为共同生活在同一群落中的物种种群处于一种稳定状态。其中心思想是:
①共同生活的物种通过竞争、捕食和互利共生等种间相互作用而互相牵制;
②生物群落具有全局稳定性特点,种间相互作用导致群落的稳定特性,在稳定状态下群落的物种组成和各种群落数量都变化不大;
③群落实际上出现的变化是由于环境的变化,即所谓的干扰所造成的,并且干扰是逐渐衰亡的。因此,平衡学说把生物群落视为存在于不断变化着的物理环境中的稳定实体。
非平衡学说的主要依据就是中度干扰理论。该学说认为,构成群落的物种始终处于变化之中,群落不能达到平衡状态,自然界的群落不存在全局稳定性,有的只是群落的抵抗性(群落抵抗外界干扰的能力)和恢复性(群落在受干扰后恢复到原来状态的能力)。
比较平衡学说和非平衡学说,除对干扰的作用强调不同以外,一个基本的区别是:平衡学说的注意焦点是系统处于平衡点的性质,而对于时间和变异性注意不足;而非平衡学说则把注意的焦点是系统在平衡点周围的行为变化过程,特别强调时间和变异性。两个学说另一重要区别是把群落视为封闭系统还是开放系统。
群落功能可从生产力、有机物质的分解和养分循环3方面来描述。
生产力群落中的绿色植物通过光合作用从无机物质制造有机化合物,这是生物群落的最重要的功能。在光合作用过程中,一段时间内由植物生产的有机物质的总量叫总初级生产力,通常以克/米2·年或千卡/米2·年表示。可是植物为了维持生存要进行呼吸作用,呼吸作用要消耗一部分光合作用生成的有机物质,剩余的部分才用于积累(生长);一段时间内植物在呼吸之后余下的有机物质的数量,叫净初级生产力,例如在森林中,60~75%的总生产量可能被植物呼吸掉,余下的40~25%才是净生产量,在水生群落中不到总生产量的一半可能被植物呼吸掉。净初级生产量随时间的前进会逐渐积累,日益增多,到任一观测时间为止积累下来的数量就是植物生物量。生物量以克/米2或千克/公顷表示。
生态学上更关心的是群落的生产力,即单位时间内的生产量。对于陆地或水底群落,是计算单位面积内的生物量数量,而对于浮游和土壤群落则按单位容积确定。因而生物生产力乃是平方米面积上(或立方米容积中)在单位时间内的生产量,经常以碳的克数或干有机物质的克数表示。
生物生产力不能与生物量混淆。例如,一年内单位面积上的浮游藻类合成的有机物质可能和高生产力的森林一样多,但因大部分被异养生物所消费,故前者的生物量只有后者的十万分之一。按照生产力,草甸草原的生物量年增长量比针叶林的大得多。根据苏联的资料,在中等草甸草原植物生物量为每公顷23吨的情况下,它们的年生产量达到每公顷10吨,而在针叶林,在植物生物量为每公顷200吨的情况下,年增长量每公倾只有6吨。小型哺乳动物比大型哺乳动物有较大的生长和繁殖速度,在相等的生物量的情况下提供较高的生产量。
消耗初级生产量的消费者也形成自己的生物量。它们在一段时间内的有机物质生产量叫次级生产量,即异养生物的生产量。消费者形成产量的速率叫次级生产力。
表1
地球上不同群落的初级生产力差别很大。R.H.惠特克按初级生产力将地球生物群落分为4类(见表1)。 绿色植物的生物生产量,一部分以枯枝落叶的形式被分解者分解,一部分被风、水或其他动力带至群落之外,一部分沿食物链传递。余下的部分以有机物质的形式积累在群落中。
有机物质的分解在许多群落中,动物从活植物组织得到的净初级生产量部分要比植物组织死亡之后被分解者细菌和真菌等利用的部分小得多。在森林中,动物食用的大约不到叶组织的10%,不到活木质组织的1%,大部分落到地面形成覆盖土壤表面的枯枝落叶层,被各种各样的土壤生物所利用。这些土壤生物包括吃死植物组织和死动物组织的食腐者,分解有机质的细菌和真菌,以及以这些生物为食的动物。虽然动物有助于枯枝落叶的破坏,但细菌和真菌在把死有机物质还原成无机最终产物方面起最主要的作用。
分解者的生物量与消费者的相比是很小的,与生产者的相比更小。然而,物质量微小的分解者的活动在群落功能中十分重要。群落中全部死亡生物的残体依赖分解者进行破坏。如果没有分解者的分解活动,生物的死亡残体将不断地积累,像在酸沼中形成泥炭那样。不仅群落的生产力可能由于养分被闭锁在死组织中而受限制,而且整个群落也将不能存在。
养分循环群落中生产者从土壤或水中吸收无机养分,如氮、磷、硫、钙、钾、镁以及其他元素,利用这些元素合成某些有机化合物,组成原生质和保持细胞执行功能。消费者动物从吃植物或其他动物取得这些元素。分解者在分解动、植物废物产品和死亡残体时,养分又释放归还到环境中,再被植物吸收。这便是养分循环,或称物质的生物性循环。例如,在森林中,某种养分从土壤被吸收进入树根,通过树的输导组织向上运输到叶子,这时可能被吃叶子的蠋所食入,然后又被吃蠋的鸟所利用,直到鸟死亡后,被分解释放归还到土壤,再被植物根重新吸收。许多养分采取较短的途径从森林树木回到土壤──随植物组织掉落到枯枝落叶层而被分解,或者在雨水淋洗下由植物表面落到土壤。
不同群落参加循环的养分数量和循环的速度不同。在一部分群落中,某些元素的较大部分保持在植物组织中,只有较小部分在土壤和水中游离。例如溶于水中的磷酸盐数量与浮游生物细胞和颗粒中的数量比较起来只是小部分。在热带森林,大部分养分保持在植物组织中,被雨水淋洗到土壤的养分和枯枝落叶腐败分解时释放出的养分,很快被重新吸收。但当一片森林被采伐或火烧后,通过侵蚀和养分在土壤水中的向下移动,造成养分的大量损失。在开阔大洋中随着浮游生物细胞和有机颗粒的下沉,养分也被携带到深处,因而在进行光合作用的光亮表层水中养分很少,所以生产力很低。4
应用群落结构在生态农业上有广泛应用,如:
1.在稻田里养鸭
2.用加工后的鸡粪喂猪
3.利用玉米螟虫天敌赤眼蜂防治虫害
4.坡上种果树 树下养家禽
5.稻藕鱼立体生态农业
本词条内容贡献者为:
李兵 - 副教授 - 西南大学
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号