科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2020-12-17
来源:中国工程院院刊
本文选自中国工程院院刊《Engineering》2020年第5期
作者:景茂峰,王源超
来源:Plant Pathogens Utilize Effectors to Hijack the Host Endoplasmic Reticulum as Part of Their Infection Strategy[J].Engineering,2020,6(5):500-504.
作为真核分泌途径中的核心细胞器,内质网介导包括钙稳态和蛋白质加工在内的多个细胞过程。病原菌在侵染植物时可以诱导内质网应激并触发未折叠蛋白反应,这一反应是一种保护性信号传导途径,在极端条件下会导致程序性细胞死亡。程序性细胞死亡是否有益于病原菌,取决于其发生的时间和方式以及病原菌是否适应于发生程序性细胞死亡的组织。
中国工程院院刊《Engineering》刊发《植物病原菌致病策略——利用效应子挟持寄主内质网》,概述了目前病原菌利用效应子挟持寄主内质网抑制植物免疫反应的这一新型致病策略。文章阐明了一些内质网蛋白被病原菌效应子所挟持来抑制内质网应激介导的植物免疫反应和相关的植物防卫反应而促进侵染的过程,有助于理解和明确植物内质网在植物与病原菌互作中的角色以及病原菌挟持和克服寄主内质网应激的分子机制,对今后制定有效的作物改良、基于激活内质网应激介导植物免疫的病虫害控制策略至关重要。
一、引言
作为真核分泌途径中的核心细胞器,内质网(endoplasmic reticulum, ER)介导包括钙稳态和蛋白质加工在内的多个细胞过程。病原菌在侵染植物时可以诱导内质网应激(ER stress)并触发未折叠蛋白反应(unfolded protein response, UPR)。作为一种保守的保护性信号传导途径,UPR在极端条件下会导致程序性细胞死亡(programmed cell death, PCD),而这可能会损害或有益于病原菌,具体取决于PCD发生的时间和方式以及病原菌是否适应于发生PCD的组织。
植物模式识别受体(pattern recognition receptor, PRR)可感知细胞表面的病原体或微生物相关分子模式(pathogen- or microbe-associated molecular pattern, PAMP or MAMP),而其生物合成和准确折叠也依赖于内质网中的N-糖基化和内质网质量控制(ER quality-control, ERQC)系统。然而,病原菌已经进化出了靶向寄主内质网应激途径介导的植物免疫(endoplasmic reticulum stressmediated immunity)的能力。最近的研究表明,病原菌将内质网应激反应作为关键的靶向目标,从而能够控制内质网应激介导的植物免疫反应。植物内质网结构也会响应病原菌的侵染信号进行重排,同时大量报道证实植物内质网传感器(ER sensor)和众多内质网蛋白参与了植物防卫相关反应(表1)。其中,一些内质网蛋白被病原菌效应子所挟持来抑制内质网应激介导的植物免疫反应和相关的植物防卫反应而促进侵染,研究阐明这些过程将有助于理解和明确植物内质网在植物与病原菌互作中的角色以及病原菌挟持和克服寄主内质网应激的分子机制。因此,本文中我们概述了目前病原菌利用效应子挟持寄主内质网抑制植物免疫反应的这一新型致病策略。
表1 植物内质网相关蛋白在病原菌侵染中的作用

GFP: green fluorescent protein; mRNA: messenger RNA; BiP: binding immunoglobulin protein; ETI: effector-triggered immunity; NTPs: NACs targeted by Phytophthora; FKBP: FK506-binding protein; PPIase: peptidyl-prolyl cis-trans isomerase.
二、靶向分子伴侣BiP抑制内质网应激介导的细胞死亡
属于HSP70(heat-shock 70 kDa protein)家族的免疫球蛋白结合蛋白BiP(binding immunoglobulin protein)是负责蛋白折叠的重要分子伴侣,它通过与内质网腔内未折叠蛋白的结合来缓和内质网应激,与内质网膜上的内质网感受器的结合来调控UPR。BiP可被非生物和生物胁迫诱导表达,并与内质网应激、PCD和植物防卫反应相关。例如,过表达本氏烟草NbBiP4 能够抑制马铃薯X病毒(potato virus X, PVX)TGBp3蛋白诱导的PCD。本氏烟草中沉默NbERD2 会减少内质网中BiP的积累,增强对内质网应激的敏感性,并加剧非寄主病原菌引起的PCD。大豆GmBiP1~4 和烟草NbBiP5 的过表达促进疫霉菌侵染并抑制BAX触发的PCD。上述研究表明BiP的积累会抑制病原菌侵染相关的内质网应激应激诱导的PCD(ER stress-induced PCD, ER-PCD)。
大豆疫霉(Phytophthora sojae)RxLR效应子 PsAvh262通过稳定BiP来增加BiP在内质网中的积累,从而抑制内质网应激介导的细胞死亡。该发现也是病原菌利用效应子操控寄主内质网应激介导植物免疫反应的首次报道。疫霉菌是半活体营养型病原菌,侵染早期在寄主中进行活体营养型生活,然后在侵染后期杀死寄主细胞进行死体营养型生活。在最初的活体营养阶段,疫霉菌利用吸器从活着的细胞组织中获取营养,因此它们通常需要有效的机制来抑制或逃避寄主PCD等防卫反应。因此,疫霉菌分泌效应子PsAvh262进入寄主细胞中利用自身的免疫球蛋白/白蛋白结合域(immunoglobulin/albumin-binding domain)来稳定BiP,阻止BiP降解,从而抑制ER-PCD。PsAvh262 沉默大豆疫霉菌失去了诱导大豆BiP积累的能力而导致ER-PCD产生、侵染能力下降。此外,BiP的过表达可以部分恢复寄主对大豆疫霉菌沉默PsAvh262转化子的敏感性,这说明PsAvh262通过抑制ER-PCD来促进侵染(图1)。此外,哺乳动物致病细菌中的几种毒素和IV型分泌系统(type IV secretion system, T4SS)效应子已通过结合BiP参与了UPR的激活和随后的炎症反应。但是,尚不清楚 PsAvh262是否参与UPR的调控。BiP在大豆和本氏烟草中的过量表达增强了对疫霉感染的敏感性,这表明BiP 负调节植物防御反应。PsAvh262也与水稻OsBiP3 相互作用,而研究表明过表达水稻BiP会显著降低XA21的积累、损害XA21介导的抗性。此外,PsAvh262 可以抑制PAMP触发的细胞死亡,这意味着由PsAvh262 介导的BiP积累可能会使受体不稳定,或者PsAvh262- BiP相互作用可能会阻止SDF2/ERdi3B/BiP复合体辅助的受体折叠,从而导致破坏下游防卫反应。
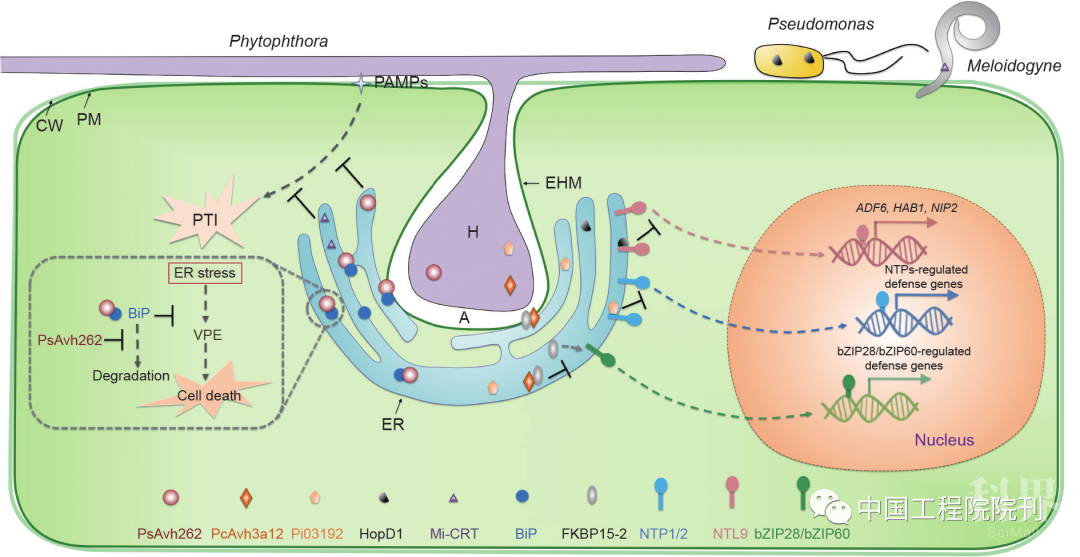
图1 植物病原菌效应子操控寄主内质网促进侵染模式图。病原菌侵染可激活寄主内质网应激反应,随后触发细胞死亡以组织病原菌的侵染和定殖。在疫霉菌侵染初期的活体营养阶段,RxLR效应子PsAvh262与寄主内质网中的BiP结合并使其稳定,从而减弱内质网应激介导的细胞死亡。RxLR效应子Pi03192阻止NAC转录因子NTP从内质网到细胞核的重定位,从而抑制NTP介导的防卫基因表达。RxLR效应子PcAvr3a12与 FKBP15-2结合并通过抑制其PPIase活性来抑制内质网应激介导的免疫反应。丁香假单胞菌III型效应子HopD1与内质网中的另一个NAC转录因子NTL9互作并抑制NTL9调控基因的表达。南方根结线虫分泌Mi-CRT进入内质网,通过抑制寄主PTI反应来促进侵染。PM:质膜(plasma membrane);CW:细胞壁(cell wall);EHM:吸器外膜(extrahaustorial membrane);A:质外体(apoplast);H:吸器(haustoria);VPE:液泡加工酶(vacuolar processing enzyme);PTI:病原物相关分子模式诱导的免疫反应(PAMP-triggered immunity);Mi-CRT:南方根结线虫-钙网蛋白(Meloidogyne incognita-calreticulin)
三、靶向内质网定位的NAC转录因子
NAC (NAM, ATAF1, 2, CUC2) 转录因子是一类植物特有的转录调节因子,在调控与应激反应相关的转录重编程过程中发挥着重要的作用。NAC转录因子通过调节活性氧(reactive oxygen species, ROS)信号传导途径、植物激素途径、内质网应激、PCD和防卫反应相关基因的表达等过程在植物防卫反应中发挥重要功能。研究证实,内质网定位的NAC转录因子在植物对调控内质网应激和抵御病原菌中起重要作用。比如,NTM1家族[NAC from the transmembrane motif 1 (NTM1)-like family]成员NTL9可以调节水杨酸(salicylic acid, SA)合成基因异丁酸合成酶1(isochorismate synthase 1, ICS1)的表达,并通过调节effector-triggered immunity(ETI)中防御相关基因的表达,在应对丁香假单胞菌(Pseudomonas syringae)的先天免疫应答中起重要作用。另外,马铃薯NAC转录因子StNTP1 和StNTP2(NAC targeted by Phytophthora)在疫霉菌培养滤液的刺激下可以从内质网中转运到细胞核中。然而,关于由NTP调控的下游基因以及NTL9或 NTP是否参与调控与内质网应激有关的下游基因了解甚少。
近期研究表明,来自丁香假单胞菌、拟南芥霜霉病菌[Hyaloperonospora arabidopsidis (Hpa)]和致病疫霉菌(Phytophthora infestans)等多种病原菌的效应子可与内质网定位的NAC转录因子互作并抑制其介导的植物防卫反应。例如,丁香假单胞菌III型效应子HopD1和Hpa RxLR效应子可以靶向AtNTL9,致病疫霉RxLR效应子Pix03192与StNTP1和StNTP2互作。III型效应子HopD1可以作为ETI的强抑制子,抑制AtNTL9调节的ETI基因表达(图1)。Pi03192可以阻止PAMP触发的 StNTP1和StNTP2由内质网向细胞核的重定位,而抑制 NTP调节的防卫反应(图1)。目前尚未发现HopD1可影响NTL9的亚细胞定位,也不清楚HopD1如何抑制依赖于NTL9的基因表达。
四、抑制内质网中FKBP的肽基脯氨酰基顺反异构酶活性
肽基脯氨酰基顺反异构酶(peptidyl-prolyl cis-trans isomerase, PPIase)催化脯氨酸肽键的顺反异构化,是蛋白质折叠过程中的重要步骤。植物中PPIase存在三个亚家族:亲环蛋白(cyclophilin, CYP)、FK506结合蛋白(FK506-binding protein, FKBP)和Parvulin蛋白,它们在植物与病原物的互作中得到了广泛的研究。例如,研究发现AtFKBP65与丁香假单胞菌和油菜黄单胞菌激发的植物防卫反应有关。最近对内质网定位的AtFKBP15-2在植物与病原菌互作中的作用进行了研究,为内质网应激与植物免疫之间的交叉提供了新的线索。结果表明AtFKBP15-2在植物感应衣霉素(tunicamycin)诱导的内质网应激、内质网应激传感器的转录表达以及UPR通路调节中起着重要作用,从而正调控内质网应激介导的植物免疫反应。
有趣的是,病原菌会招募植物CYP蛋白ROC1和 GmCYP1,利用其PPIase活性来帮助自身效应子的激活以促进病原菌的侵染。辣椒疫霉(Phytophthora capsici)中RxLR效应子Avr3a家族成员PcAvr3a12,可以与AtFKBP15-2互作,通过抑制AtFKBP15-2的PPIase 活性而减弱其介导的免疫反应(图1)。同时,AtFKBP15-2的PPIase活性在其调控的内质网应激介导的免疫反应中起着关键作用。
五、分泌CRT效应子进入寄主细胞
钙网蛋白(calreticulin, CRT)是在内质网腔中高度保守的钙结合分子伴侣和钙离子传感器。CRT与钙离子稳态有关,并参与了植物的蛋白质折叠和植物免疫反应。CRT3在膜定位受体EFR、BRI1和IRK的积累中具有重要作用。拟南芥crt3突变体对丁香假单胞菌的感病性增加,然而过表达CRT2 不仅可以增加寄主对丁香假单胞菌的感病性,也能导致水杨酸的积累和病程相关基因的激活,表明了CRT家族蛋白在植物免疫调节中的双重作用。南方根结线虫(Meloidogyne incognita)在侵染寄主过程中,会分泌CRT (Mi-CRT) 定位到寄主内质网和高尔基体中。而研究发现多种动物寄生虫也会分泌CRT到它们的宿主中,并在调节宿主防卫反应中发挥重要作用。在南方根结线虫中敲除Mi-CRT会导致在侵染番茄和拟南芥时毒性降低,这表明了Mi-CRT对于线虫成功侵染寄主的重要性。同时,在寄主中过表达Mi-CRT不仅增加了植物对南方根结线虫的敏感性,还促进了病原卵菌寄生疫霉菌(Phytophthora parasitica)的侵染。Mi-CRT通过抑制 PAMP触发的免疫(PAMP-triggered immunity, PTI)和植物激素介导的防卫反应而增加了植物对病原菌的感病性。该研究提供了一个抑制寄主免疫反应的例子,即病原物模拟并分泌寄主内质网应激相关途径的分子伴侣蛋白。
六、总结
总之,病原菌与植物内质网应激的互作机制研究处于起步阶段,还有诸多关键的科学问题需要研究,比如:植物如何识别内质网应激信号并调节其介导的植物免疫?在植物与病原菌互作中,植物如何平衡内质网应激介导的自噬与细胞死亡?病原菌如何将效应子分泌到寄主内质网中,而效应子又是如何发挥作用的?仍有一系列的问题需要进一步的研究和探索。此外,深入理解植物对病原菌诱导的内质网应激信号的识别、病原菌如何突破内质网应激介导的植物免疫而促进病原成功定殖,对于制定有效的作物改良、基于激活内质网应激介导植物免疫的病虫害控制策略至关重要。
注:本文内容呈现略有调整,若需可查看原文。
改编原文:
Maofeng Jing, Yuanchao Wang.Plant Pathogens Utilize Effectors to Hijack the Host Endoplasmic Reticulum as Part of Their Infection Strategy[J].Engineering,2020,6(5):500-504.
来源:CAE-Engineering 中国工程院院刊
原文链接:http://mp.weixin.qq.com/s?__biz=MzA3MTUwNjU3NQ==&mid=2651038660&idx=3&sn=01525dcd9c167b04c8bb6dfa824fc57b
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn
第二届“药食同源与植物代谢”国际学术研讨会在沪举行
植物生理生态研究所科研人员发现植物抗虫调控新机制
李新荣:沙漠书写草根论文
《生命科学》出版植物营养与人类健康专刊

盘点影响人类生活的十大植物病原菌
2017国际植物日之--南开大学站成功举办

30秒!人类、动植物疾病及病原菌检测,新试纸全搞定

首次发现!病原菌分泌细胞壁降解酶,帮助感染植物
中国植物生理与植物分子生物学学会植物生物学女科学家分会第三届学术交流会成功举办
Microbiome:根系分泌物驱动土壤记忆抵御植物病原菌


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号