科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2021-02-03
来源:BioArt植物
植物主要依靠细胞膜上的免疫受体(PRR)识别病原菌的微生物相关分子模式(MAMP,如细菌鞭毛flg22和真菌几丁质)激活模式触发式免疫(PTI),从而获得对病原菌的基础抗性。PTI的持续激活会抑制植物生长。因此,PTI的负调控因子具有非常重要的生理功能。另一方面,病原菌能够分泌致病因子进入植物细胞,通过负调控PTI而造成植物的感病性(ETS)。进而,植物能够利用胞内的免疫受体(R蛋白)感知致病因子或其破坏效应,激活以细胞死亡为主要形式的效应触发式免疫(ETI)。上述即为经典的植物-病原菌互作的“Z”字模型。举一例如下:
拟南芥PTI的负调控因子RIN4可结合FLS2并抑制其激活,以防止FLS2在未识别flg22的情况下激活免疫;FLS2对flg22的识别能导致RIN4的Ser141位点发生磷酸化,解除对FLS2的抑制而激活PTI;丁香假单胞菌分泌的致病因子AvrRpm1能够操纵未知的拟南芥激酶,磷酸化RIN4的Thr166位点,从而抑制flg22诱导的Ser141磷酸化,使RIN4保持对FLS2的抑制而造成ETS;拟南芥进而通过R蛋白RPM1识别RIN4的Thr166磷酸化,激活ETI。然而,AvrRpm1究竟利用了什么宿主激酶使RIN4的Thr166位点发生磷酸化一直有待阐明。
2021年2月2日,中山大学李剑峰课题组在The Plant Cell发表了题为The receptor-like cytoplasmic kinase CDG1 negatively regulates Arabidopsis pattern-triggered immunity and is involved in AvrRpm1-induced RIN4 Phosphorylation的研究论文,揭示了拟南芥类受体胞质激酶CDG1是PTI的负调控因子,而AvrRpm1能够诱导CDG1磷酸化RIN4的Thr166位点,造成植物的ETS。


在该研究中,作者首先通过在拟南芥原生质体中逐个过表达23个类受体胞质激酶(RLCK)VII家族成员,发现过表达CDG1能够抑制flg22引发的MAPK激活。随后构建了CDG1过表达或敲除植株,从植株水平确认了CDG1对flg22信号转导的负调控作用,同时发现CDG1也参与了对几丁质信号转导的负调控。接下来,作者发现CDG1能够与flg22受体FLS2和几丁质共受体CERK1互作,而CDG1过表达引起的自激活能够造成FLS2和CERK1的降解。进一步研究发现,CDG1还能与MAPK级联通路中的 MEKK1(MAPKKK8)发生互作。意外的是,在MEKK1激酶失活背景下,CDG1过表达则不会引发FLS2和CERK1的降解;反之,雌二醇诱导的MEKK1过表达或组成型激活的MPK4 D198G/E202A突变体均能促进FLS2降解。这些结果表明CDG1及其下游MEKK1介导的MPK4通路参与了对flg22激活的FLS2稳定性的负反馈调控。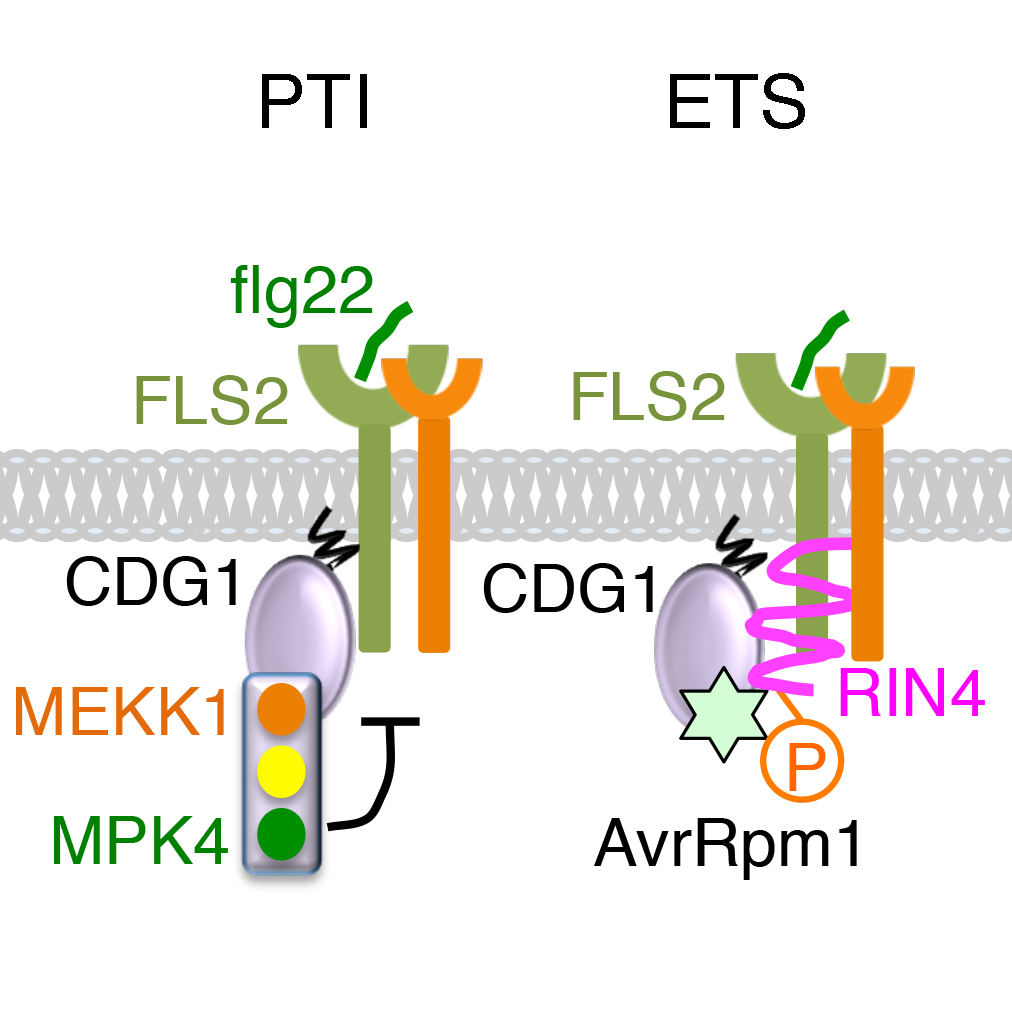 另一方面,作者发现AvrRpm1能够诱导CDG1与RIN4发生互作并在体外磷酸化RIN4的Thr166位点。在cdg1突变体中,AvrRpm1诱导的RIN4 Thr166磷酸化以及该磷酸化触发的细胞死亡都显著减弱,提示CDG1是被AvrRpm1操纵并磷酸化RIN4 Thr166位点的宿主激酶之一。最后,作者发现AvrRpm1表达也能够诱导FLS2的降解。该降解完全依赖于RIN4,部分依赖于CDG1,但不依赖于MEKK1的激酶活性,说明PTI中CDG1调控的FLS2降解与ETS中AvRpm1调控的FLS2降解经由不同机制。
另一方面,作者发现AvrRpm1能够诱导CDG1与RIN4发生互作并在体外磷酸化RIN4的Thr166位点。在cdg1突变体中,AvrRpm1诱导的RIN4 Thr166磷酸化以及该磷酸化触发的细胞死亡都显著减弱,提示CDG1是被AvrRpm1操纵并磷酸化RIN4 Thr166位点的宿主激酶之一。最后,作者发现AvrRpm1表达也能够诱导FLS2的降解。该降解完全依赖于RIN4,部分依赖于CDG1,但不依赖于MEKK1的激酶活性,说明PTI中CDG1调控的FLS2降解与ETS中AvRpm1调控的FLS2降解经由不同机制。
中山大学博士生杨秋姣为论文的第一作者,李剑峰教授为唯一通讯作者。该研究得到国家自然科学基金委面上项目的资助。
原文链接:http://mp.weixin.qq.com/s?__biz=MzU3ODY3MDM0NA==&mid=2247502651&idx=2&sn=cc08e00996b67dd201153a58ee4b9c6a
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn
蛋白质酪氨酸磷酸化的分子识别研究
第二届“药食同源与植物代谢”国际学术研讨会在沪举行
第二届“分子植物”国际学术研讨会在清华大学成功召开
综述:生长素途径中的蛋白可逆磷酸化机制

Nat Biotech:“机器学习”帮助鉴定磷酸化位点
2015上海辰山“药食同源与植物代谢”国际学术研讨会顺利召开
中国植物生理与植物分子生物学学会植物生物学女科学家分会第三届学术交流会成功举办
《生命科学》出版植物营养与人类健康专刊
2017国际植物日之--南开大学站成功举办

植物所揭示糖基化和磷酸化修饰介导小麦开花的新机制


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号