科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2021-02-23
来源:BioArt植物
2020年2月22日,美国冷泉港实验室 (CSHL) David Jackson研究组在Nature Plants发表了题为Enhancing grain-yield-related traits by CRISPR–Cas9 promoter editing of maize CLE genes的论文(刘磊博士为该论文的第一作者)。该研究利用CRISPR/Cas9系统对玉米CLE基因启动子进行编辑,创制了玉米高产等位基因。

作物产量的遗传改良育种,经历了数次变革。第一代的育种策略,是基于种质源收集、评估和筛选,保留高产优质种质资源进行种植;第二代育种策略标志性事件是杂交育种概念的提出和应用,通过自交系间杂交,获得杂交种,而杂家种由于杂种优势的效应,其产量显著高于双亲,同时化肥杀虫剂等应用,以及矮秆基因在多个作物中的应用,提高了作物的产量;随着生物技术的发展,通过转基因技术、分子标记辅助选择、全基因组选择等,进一步提升了作物的产量等。然而,随着全球人口激增,气候变化对作物产量的不利影响,对基于玉米等作物的生物能源的需求增长等,对作物产量提出了更高的要求。现代玉米的驯化和改良起源于约10,000年前,由玉米野生种大刍草 (Zea mays ssp. Parviglumis) 经过长期的驯化选择,演化为玉米地方品种(landrace),再经过约150年的遗传改良,进而发展出适应性好、籽粒产量高的优良玉米品系。在此过程中玉米对光周期适应性、植株形态、果穗大小、籽粒产量等性状,发生了极为显著的变化(Duvick 2005; Doebley et al 2006)。其中,玉米的产量性状受到强烈的选择,野生种大刍草果穗仅有数十粒籽粒,而现代玉米果穗则多达数百粒,提升玉米单穗的籽粒数可显著提升玉米的产量。
玉米是雌雄异花植物,玉米的果穗(雌穗)为雌花序,发育出雌花,并通过接受雄穗雄花的花粉,受精后最终发育成籽粒。玉米果穗籽粒数目,是在受雌花序发育早期形成和决定的(Vollbrecht and Schmidt 2009)。玉米的雌花花序一般在9-10叶期开始发育,此时位于叶腋处的叶腋分生组织(Axillary Mersitem, AM)转换成花序分生组织(Inflorescence Meristem, IM)(Vollbrecht and Schmidt 2009)。随后,IM上形成多行排列整齐的分生组织,即成对小穗分生组织(Spikelet-Pair Meristem, SPM)。每一个SPM进而分化出两个短分枝的分生组织:小穗分生组织(Spikelet Meristem, SMs)。每个SM最终经历小花分生组织和花器官的分化,授粉后发育成成熟的籽粒(Vollbrecht and Schmidt 2009)。因此,玉米雌穗发育过程中IM持续分化SPM的能力及SPM分化SM的能力,决定了最终玉米果穗上的每穗行数、行粒数等产量性状(Vollbrecht and Schmidt 2009)。因此,提升玉米花序分生组织活性,理论上可增加果穗籽粒数目,提升产量。
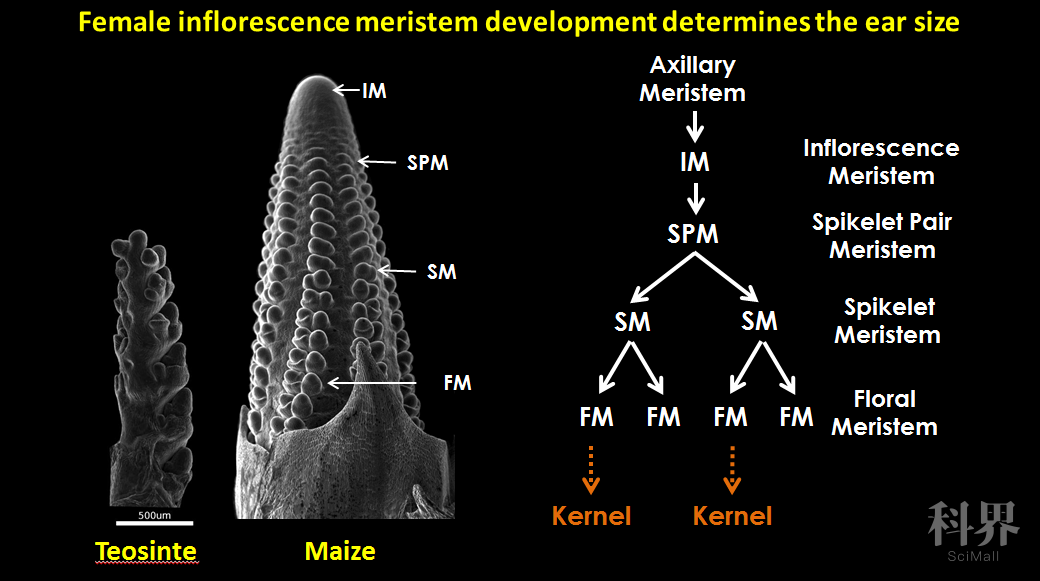
玉米花序分生组织(IM)的建成和维系,是玉米花序正常发育的基础。和其他分生组织类似,IM主要发挥两方面作用:通过植物干细胞(Stem cell)的分化起始其他组织器官的发育;通过细胞的分裂进行自身的增殖(Vollbrecht and Schmidt 2009)。在植物中,维持花序分生组织生长和分化的经典途径是CLAVATA-WUSCHEL feedback loop,其中包括在IM中部特异表达的WUSCHEL(WUS)基因,在IM中特异表达的CLAVATA1(CLV1)和CLAVATA2(CLV2),以及负责WUS向CLV1和CLV2进行信号传递的CLAVATA3(CLV3)信号分子(Williams and Fletcher 2005)。拟南芥中WUS基因的产物可促进分生组织的生长及CLV3信号分子的表达,CLV3可以和CLV1-CLV2复合体互作,进一步抑制WUS的表达,因此形成一个动态平衡的负反馈回环通路,维持分生组织的分化活性(Williams and Fletcher 2005),打破此通路平衡的突变体,表现为分生组织过度增殖(Williams and Fletcher 2005)。CLV-WUS通路在植物中相对保守,在玉米中鉴定了拟南芥CLV1、CLV2和CLV3的同源基因突变体thick tassel dwarf1(td1)(Bommert et al 2005)、fasciated ear2(fea2)(Taguchi-Shiobara et al 2001; Bommert et al 2013a)及Zmcle7、Zmfcp1(Rodriguez-Leal et al 2019)。这些突变表型类似,均表现为IM的增大,雄穗雌穗增粗、穗行数增加等表型。然而这些突变体,由于分生组织过度增殖,导致果穗发育异常,因而显著降低产量。美国冷泉港实验室David Jackson研究组利用EMS等技术创制的fea2的弱突变体,可保证果穗正常发育,且提升了分生组织活性,进而数量上提升了穗行数等产量性状,因此通过对这些基因的人为改造,精细优化它们的生物学活性,对优化玉米分生组织活性、提升玉米产量,具有重要的潜在价值。Rodríguez-Leal等人利用CRISPR-Cas基因编辑技术,对番茄的CLV3基因进行调控序列的编辑,获得了可以调控番茄果实大小数量变异的新的等位基因,实现了利用基因组编辑技术对目的基因进行人为设计(Rodríguez-Leal et al 2017)。因此应用基因组编辑技术可以进一步创造优良等位基因资源,快速应用于玉米的遗传改良,在玉米育种中的具有广泛的应用前景(Wolter et al. 2019)。
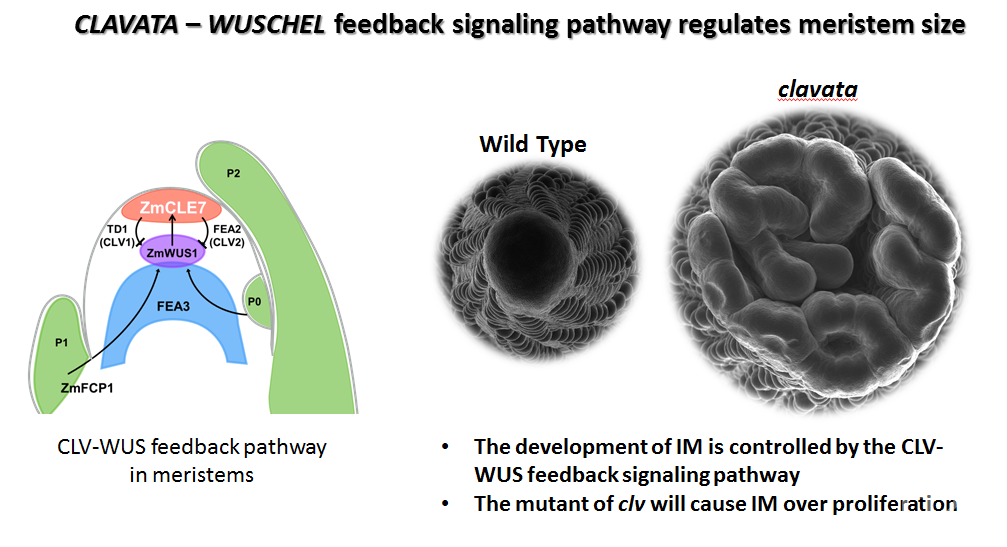
玉米的CLAVATA3(ZmCLE7和ZmFCP1)是一类短肽信号分子,其将信号传递给CLAVATA1(TD1)、CLAVATA2(FEA2)及FEA3,并通过他们抑制WUSHEL基因的表达,进而形成一个反馈调控的信号通路,精准调控花序分生组织发育。为了获得其弱突变等位基因,该项最新研究探索利用CRISPR-Cas9基因组编辑技术,靶向其启动子区域,获得影响其表达量的弱突变体。针对ZmCLE7和ZmFCP1启动子区域,研究者利用ATAC-seq和MNase-seq等数据进行染色质开放区域(accessible chromatin regions)的分析,同时利用进化分析,预测启动子区域可能的保守的调控位点,并分别设计9个sgRNA,靶向其启动子区域。通过筛选,ZmCLE7和ZmFCP1启动子区域分别筛选到数个大片段的缺失和倒位的编辑事件。这些编辑事件可显著改变基因的表达水平,数个启动子编辑等位基因可使表达量数量水平降低~45% - ~69%。其中ZmCLE7启动子倒位编辑的等位基因,却使得ZmCLE7表达部位扩展到IM内部。
通过进一步的表型分析发现,一些启动子缺失编辑的等位基因,可一定程度的提升花序分生组织的大小,进而显著提升数个产量相关性状及单穗的籽粒产量。它们在自交系背景及杂交种背景下,均能显著提升产量。这些等位基因对产量的提升,主要体现在果穗增粗、穗行数增加、单穗粒数的增加;相对于对照,在穗行数上的提升十分显著,提升了~6行,且粒重未出现显著变化,最终单穗产量提升约最多至~26%。该研究还发现,其中一个启动子倒位的编辑等位基因,扩张了ZmCLE7的表达部位,因此导致果穗变小,降低了产量。这些结果表明,对ZmCLE7和ZmFCP1启动子的编辑,可以通过数量上调节基因表达,影响其对玉米花序分生组织活性的影响,影响玉米产量性状的数量变异,显著提升玉米的单穗产量。
玉米中的CLAVATA3(ZmCLE7和ZmFCP1)的同源基因共有49个,研究者前期研究发现,CLAVATA3及同源基因,在多物种中往往都具有功能冗余的特性。例如在Zmcle7突变体中,ZmFCP1可通过表达量上调,弥补一定的ZmCLE7的功能。研究者进而通过对Zmcle7突变体转录组的分析,筛选到了另外一个CLAVATA3同源基因ZmCLE1E5,其在Zmcle7突变体中,也表现为表达量上调。研究者对其编码区进进行编辑,获得了两个不同移码突变的null alleles突变体。Zmcle1e5突变体的花序分生组织发育相对正常,但是显著增大,果穗显著增大,穗粒数增加,果穗形态正常,产量显著提升,且Zmcle1e5可显著增强Zmcle7突变表型,因此,ZmCLE1E5可部分互补ZmCLE7的功能。进而表明,同过对ZmCLE7互补因子的基因编辑,同样可以创制ZmCLE7启动子编辑类似的高产等位基因。
玉米产量是极为复杂的数量性状,前人利用全基因组关联分析、连锁分析等手段,鉴定了数百个产量相关性状的QTL位点,其中大部分为微效QTL位点,且仅有少数位点被克隆,例如穗行数QTL位点KRN4,控制百粒重的QTL qHKW1、控制行粒数的KNR6等(Liu et al 2015; Yang et al 2019; Jia et al 2020)。这些QTL位点往往仅在一定的遗传背景下,才表现出较大的遗传效应,例如KRN4位点在H21背景下,可增加穗行数2.2行,而在Mo17背景下,其穗行数效应只有0.7行(Liu et al 2015)。另外,这些QTL位点的优良等位基因,往往在优良自交系群体中已被富集,进一步限制了这些位点在育种中的应用。而另外一方面,一些控制产量相关性状发育的关键基因,却不存在可导致表型数量变化的自然变异位点,一方面可能由于这些基因的优良自然变异位点本身就不存在,或者在驯化和改良中由于瓶颈效应丢失。例如,本研究涉及的两个CLAVATA3同源基因ZmCLE7和ZmFCP1,在具有广泛遗传变异的NAM群体里,并未检测到控制相关性状的QTL或关联位点(Brown et al 2011)。因此,该研究利用基因组编辑技术创制的ZmCLE7和ZmFCP1优良等位基因为新的基因资源。ZmCLE7和ZmFCP1调控区域编辑的优良等位基因,精细调控候选基因的表达水平,平衡其在花序发育中的功能,最终强化玉米产量性状表现,数量水平提高玉米籽粒产量,且它们的产量相关性状的效应,大于目前已克隆的任意QTL位点(Liu et al 2015; Yang et al 2019; Jia et al 2020)。除了启动子编辑策略,该研究还提出了对分生组织发育关键基因的互补基因进行编辑,也可优化分生组织活性,创制高产优良等位基因的策略。综上所述,该研究一方面创制了优良等位基因资源,另一方面,为重要基因的再利用提供了新的思路。
来源:bioartplants BioArt植物
原文链接:http://mp.weixin.qq.com/s?__biz=MzU3ODY3MDM0NA==&mid=2247503348&idx=1&sn=208d7de509c1616e2abc1d8f437b914d
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn

基因编辑技术让奶牛不长角

神奇的剪刀:基因编辑技术!
剪掉“坏基因”,“上帝的手术刀”难免失手

基因编辑技术育出节水番茄

昆虫基因编辑获突破,新技术首次实现对蟑螂基因编辑
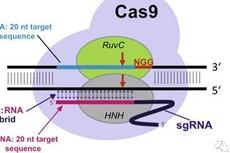
CRISPR/Cas9基因编辑技术
基因编辑介导的基因驱动技术助力植物快速改良
一图了解基因编辑技术
赵云德:无转基因残留的基因编辑技术

巴西培育基因编辑“供体猪”


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号