科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2021-04-28
自我的面孔对人类来说有着重要的意义,它代表着一个人的身份以及自我感知。相应的,面孔也有着认知上的优势,大脑处理自我面孔的速度要比他人面孔信息更快(即使是和很熟悉的他人面孔相比)。自我面孔的特殊性不仅仅体现在行为层面,fMRI成像研究报告了一些脑区对自我面孔的反应要比他人面孔强。并且,当一个人的面孔由于化妆或者P图显得更好看时,会激活多巴胺奖励通路,例如腹侧被盖区(ventral tegmental area,VTA)还有伏核(nucleus accumbens,NA),但他人面孔就没有出现这种效应。相反,和恐惧情绪相关的杏仁核则对他人面孔反应更强。有趣的是,有时候这种现象的发生甚至是无意识的。
Ota和Nakano设计了一个小实验来探究人们在无意识的情况下对不同种类面孔的识别机制。实验包括22名20-25岁的女性被试,在fMRI扫描之前研究者给每位被试的面孔拍10张照片(无情绪;分别为正脸,发“a”,”i”,”u”,“e”,“o”,以及脸向左、右、上、下分别倾斜)。另外研究者还采集了13名其它女性(20-25岁)的不同表情的照片。接着研究者给这些照片美颜:放大眼睛和瞳孔,并且为他们瘦脸,程度分为轻微和极端两种水平(Fig. 1B)。轻微美颜处理是通过将眼睛放大1.1倍,瞳孔放大1.1倍,将下半脸瘦至原来的0.9倍;极端美颜是通过将眼睛放大1.21倍,瞳孔放大1.21倍,瘦脸至0.81倍。
站在面前的是小哥哥还是小姐姐?
我要场内求助mPFC!
你怎么知道路边的小花是只猫,教室里的大黄是只狗?学习!其中一个非常重要的环节——归类总结(categorisation)可不就是成为学霸的必胜法宝嘛。只要在学习中多领悟多归纳,那么成千上万的考题在你眼里最终不也就是那么几个老生常谈的课后练习题。日常生活中,我们能精确地判断一个动物是狗还是猫,也依赖于大脑对学习过的内容进行了妥善的管理分类。对某一类而非某一个事物的记忆,有利于我们变通行为选择,这对我们在复杂社会中生存至关重要。已有研究表明,灵长类动物的大脑皮层,特别是前额皮质(prefrontal cortex,PFC)中的神经元可以灵活地表现出类别选择性。然而,这些神经元的代表性是如何首次在动物大脑中出现的呢?是日复一日的学习训练使之逐渐成为语义记忆的一部分,还是任务执行中被立即分配到不同的小组呢?
近日,来自德国马克斯·普朗克神经生物学研究所的Goltstein研究组首次揭露了,小鼠前额皮质的类别表征是在学习过程中逐渐获得的,它们通过不断学习并归纳各类刺激来总结出相应的规则,并以此类推到新来的刺激以便做出更合适的选择。
研究人员首先利用基于“走/不走”(‘Go’/‘NoGo’)的分类任务来训练老鼠,让它们学会将所接触的多种刺激根据相应的规则分成两类。这些刺激都包含这两个特征信息:空间频率和方向。但要注意的是训练只会由其中的某一种特性来决定分类规则(有效规则),比如说系统只会根据视觉刺激的方向来分配它们的类别身份(即使他们的空间频率不同,他们也会被视为同一类刺激)。小鼠可以在短期训练后将新的刺激总结归纳到前面已经学会的分类中,准确的知道这个信号代表的是走还是不走。因为“规则转换”(rule-switching)学习在分类训练中尤为重要,已经接受过第一个规则训练(比如方向)的小鼠需要在新的训练中将同样的刺激根据第二种规则(空间频率)进行重新分类。小鼠通过对18个刺激信号进行分类学习后,能很快地根据新的分类规则准确类推到新的刺激中。这说明小鼠能够基于规则区分不同类别的刺激。利用这个训练模式,研究者们接着就想进一步了解分类学习中神经元之间的秘密。
视力再差,
也绝不让视皮层背锅
小鼠是出了名的近视眼,作为神经领域的头号明星选手,它初级视皮层的神经细胞数量只有灵长类的1/1500,就算进了卢浮宫,再美丽的蒙娜丽莎在它看来也只是一团马赛克般的“朦胧丽莎”。
很长时间以来,大家都认为是视觉神经细胞的灵敏度低导致了小鼠的先天近视,真是人在家中坐,锅从天上来。据来自霍华德·休斯医学研究所(HHMI)Janelia Research Campus的Marius Pachitariu团队,小鼠的视皮层细胞终于能沉冤昭雪:这些神经细胞能区分0.35度的视觉差别,这比它实际行为上的分辨能力高了100倍!所以,视皮层以后终于不用做背锅侠啦。
研究人员的初衷是想知道少数几个神经元的噪声是否会对整体视觉编码的准确性造成影响,为此,他们利用多层双光子钙信号成像技术(multi-plane two-photon calcium imaging),记录了约五万个细胞在视觉分辨任务中的发放情况。
研究人员发现单个细胞对视觉刺激的放电波动很大(信噪比低达0.13)。由此推测,单个细胞的信号噪声可能以三种方式影响整体视觉编码的准确性:互相独立的(independent),部分相关的(partially correlated),或者完全相关(fully correlated)。如果细胞的信号波动彼此相互独立的话,那么理论上研究人员可以用朴素的贝叶斯分类器(naïve Bayes classifier)来解码细胞群体编码的视觉方位信息,结果他们用贝叶斯解码的方法得出的中位数误差为2.31度——一个不大也不小的数。那么,这个误差到底是来自单个细胞的误差,还是由细胞之间解码误差的高度相关造成的呢?研究者进一步将细胞分为两份,并分别用同样的方法分类以后再相互比较,他们发现两个误差相对于真实值呈现相同的误差趋势,并且高度相关,这说明用朴素贝叶斯分类中细胞波动相互独立的前提不成立,所以第一种相互独立的假说并不成立。
上一个实验结论也说明,如果考虑到细胞波动的相关性问题,解码误差将可能进一步降低。因此,研究者继而换用了另一个简单的线性分类器来解码视觉信号的方位。他们训练了一些介于所有细胞和最终解码结果之间的“超级神经元(super-neurons)”,这些超级神经元对不同的角度有各自的选择性,能综合考虑每一个细胞的权重,特别是能降低对解码具有毁灭性打击的细胞权重。研究者发现,这种线性分类器能将解码的中位误差降低到1.03度,大大低于之前的2.31度。不仅如此,如果不断增加神经元数量,或者继续增加刺激试验的次数,这一方法的解码误差还能进一步被降低。事实上,研究人员将试验次数从每一度10次提高到了每一度1000次,结果发现甚至这些细胞的信息能分辨相差低于0.1度的两种刺激!
决策脑:最后关头愈加坚定
我们的各路感官每时每刻都在经历信息轰炸,而决策则需要大脑从一团乱麻中梳理头绪——只对感兴趣的输入做出正确的反应。大脑是如何排除干扰物对决策的影响的呢?
Finkelstein等人设计实验采用光遗传方法刺激小鼠的vS1(vibrissal somatosensory cortex;Fig. 1a)的layer4神经元,同时采用硅探针记录vS1和ALM(anterior lateral motor cortex,该脑区参与控制定向的舔舐动作;Fig. 1d,e)脑区锥体神经元的活动。实验分为sample和delay两个阶段,由声音提示区分。在实验中,小鼠需要学会的任务是:在sample阶段,如果研究者刺激小鼠的vS1,那么小鼠需要在delay阶段舌头舔向右边;而如果没有接收刺激,则小鼠需要舌头舔向左边(Fig. 1b,c)。这样的实验设计可以将感觉输入信息和决策信息表征很好地分开。
实验中55%(300/547)的vS1神经元活动受到了光遗传学刺激的调制,vS1神经元在光刺激过程中达到峰值随后反应迅速降低(Fig. 1f);而仅有13%(316/2417)的ALM神经元片段性地对光刺激反应(Fig. 1e,示例细胞5),这部分细胞多位于与所刺激的vS1同侧半脑,而双侧半脑的大多数神经元在delay时间段活动缓缓提高(Fig. 1e,示例细胞6和7表现出对实验条件的选择性,示例细胞8无选择性)。
Finkelstein等人采用两个脑区的神经活动来解码感觉输入(是否光刺激)和行为决策(舔舐的方向),如Fig. 1h所示,vS1神经元对感觉信息的解码先达到峰值,而几乎完全不编码决策信息,而ALM神经元活动对决策信息的解码准确度则在delay时段内逐渐升高。以上的结果表明,光刺激能够对两个脑区的神经元活动都产生影响,其中ALM神经元在delay时段会对下来的决策编码。

Fig. 1 | 采用光刺激输入运动感觉信息
Finkelstein et al., Nat Neurosci.
在小鼠学会基本任务以后,Finkelstein等人接下来在实验中加入干扰:在小鼠舌头舔向左边(也就是此前未受到光刺激)的那些试次中的不同时间点施加弱的光刺激干扰(Fig. 2a),这个实验中用到的小鼠此前在训练中从未遇到过这种情况。
研究者进而发现,如果在sample阶段或者delay阶段的早期施加干扰,小鼠随后的行为会受到影响(即舌头更多地舔向右边),而如果在delay阶段晚期施加干扰则不会影响行为,这一现象表明出现了时间门控效应(Fig. 2c左)。而如果在delay晚期施加的是强干扰,则还是会影响动物行为(Fig. 2d左),也就是说门控效应除了取决于时间,还取决于干扰强度。研究者另外训练了一批小鼠,在训练阶段就采用不同强度的干扰,但如果在受到干扰后选择舔向右边则不会得到奖励。在这种情况下,delay阶段早期或晚期的弱干扰都不会显著改变小鼠的行为(Fig. 2c右),但强干扰项的影响仍然存在。
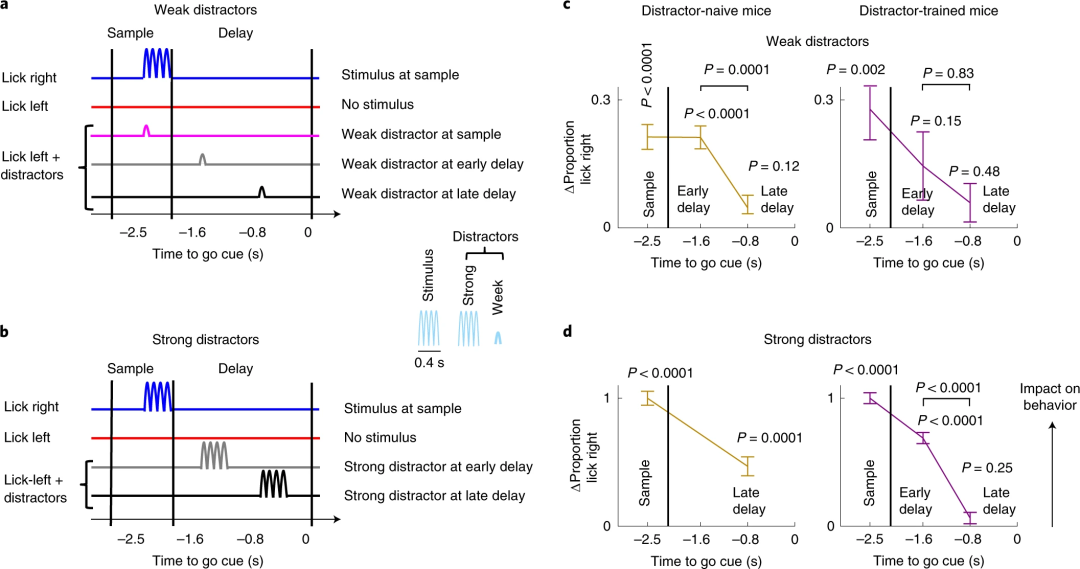
Fig. 2 | 在决策中对感觉输入的门控效应
Finkelstein et al., Nat Neurosci.
对以上的现象Finkelstein等人提出两种可能的假设:(1)当在delay阶段后期引入干扰时,刺激和干扰触发的vS1或是ALM神经元反应被抑制(干扰抑制;Fig. 3a);(2)在delay阶段刺激输入信号都能到达ALM,但在越接近要发生动作的时候,神经网络活动让运动计划更鲁棒(Fig. 3b),也就是说感觉输入的门控其实发生在行为层面。
进一步的分析并未在vS1观察到任何干扰抑制的现象(Fig. 3c),说明vS1并不存在门控。而对ALM的分析同样表明干扰信息在到达ALM后并未减弱(Fig. 3d),但干扰的确对行为产生了影响。由于ALM脑区和刺激、决策相关的信息处理都相关(Fig. 1h),研究者对ALM神经元活动进行降维处理,将刺激相关和决策相关信息分离开来(Fig. 3e),结果并未发现刺激相关的干扰抑制现象(Fig. 3f),也就排除了第一种假设。而对决策相关的神经反应活动进行解码,发现和随后发生的行为吻合度很高。
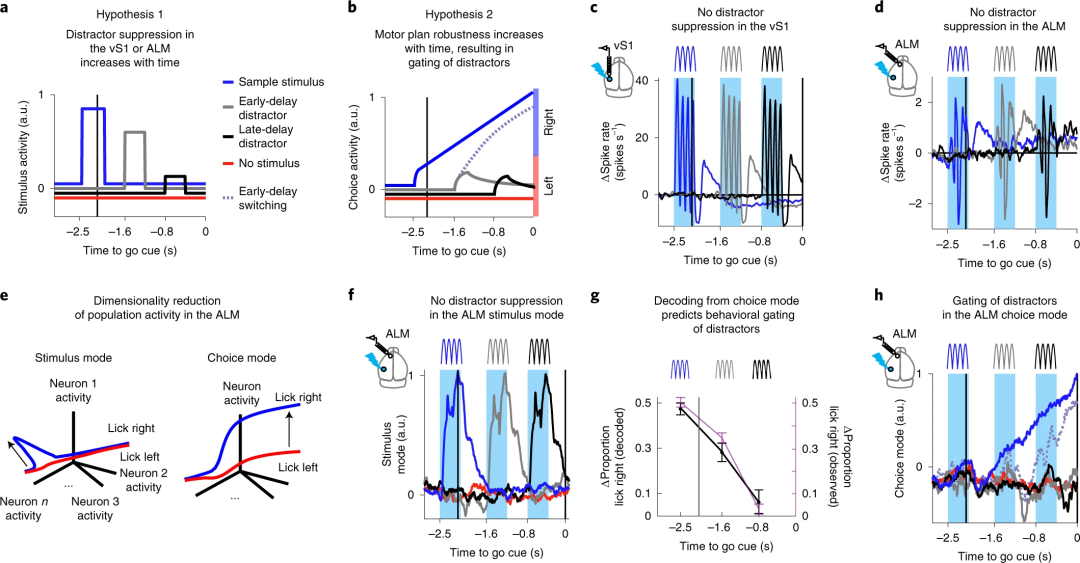
Fig. 3 | ALM群体反应未表现出干扰抑制
Finkelstein et al., Nat Neurosci.
特别的,对于那些中途受到干扰但被动物忽略的试次中,可以观察到ALM与决策相关的活动首先产生扰动又还原(Fig. 3h)。这一结果说明那些最终行为没有被干扰项影响的试次中,实际上决策相关的活动还是受到了干扰项的感觉输入信息的影响。相反,那些在delay阶段早期受到干扰最终影响行为的试次中,ALM神经活动也反映了这一变化。总的来说,以上的结果更加支持第二种假设。
在决策中面对各种干扰,大脑需要一道闸门。一般研究者们认为在这个过程中,被注意的感觉刺激的神经反应会增强,反之无关刺激(不被注意的干扰刺激)的神经反应会减弱。对于这种神经门控过程背后的神经网络层面的机制,一直以来研究者们提出了很多假设:新皮层自上而下的调控、丘脑的增益作用、兴奋和抑制作用的平衡(一些输入的信息可能被局部的抑制抵消)、由不同脑区的神经元群体反应同步性来选择性传递信息,等等。这些对门控机制都隐含着一个假设,那就是当输入信息从一群神经元传递到另一群神经元时,相对感兴趣刺激的神经反应来说,对干扰刺激的神经反应应该被抑制。而Finkelstein等人报告的结果说明,这道闸门并不是设在感觉输入通道上;实际上是决策相关的神经环路活动调控了最终的行为,而且在越是接近要发生行为的时刻,决策相关的神经表征就越是鲁棒。
来源:神经现实
原文链接:http://mp.weixin.qq.com/s?__biz=MzI0MjI1NTgxNQ==&mid=2651441952&idx=1&sn=5366bac34d6acbc7f63604c5b2aed653
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn
脑损伤激活胶质细胞产生神经元研究获进展
简单小分子组合实现胶质细胞直接转化为神经元
脑智卓越中心小鼠皮层神经元细胞群体同步激活研究获进展
小胶质细胞监测和保护神经元的功能
大脑“后勤”细胞参与指挥神经元发育
揭秘癌细胞与神经元的“共生共舞”

经典论文:癌细胞的行为类似于神经元

人类大脑有多少个神经元细胞

高盐饮食引发小鼠大脑认知功能缺陷

人类皮肤细胞直接“变身”运动神经元


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号