科技工作者之家
科界APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2020-04-10
来源:brainnews
Theta节律是一种4-8Hz震荡,最早于上个世纪30年代在动物大脑中被发现。在多种自发性的活动中,例如运动准备,朝向,觅食,甚至在非运动状态(清醒)和睡眠过程中,我们都可以在啮齿类动物的大脑上记录到theta活动。近几十年来,大量研究表明theta震荡在记忆的编码和提取中有着重要作用。最早的一项将theta活动和记忆联系起来的研究证据来自于大鼠。该研究发现,给大鼠的足部进行电刺激后,其大脑的theta节律的持续时间和足部刺激记忆保持的程度呈正相关【1】。之后大量的研究进一步提供了theta节律和记忆的证据,尤其是海马中的theta的能量,相位和情景记忆,空间记忆密切相关。
海马中的theta产生的生理机制
已有研究发现theta活动在海马最为明显,在海马子区CA1,CA3以及齿状回都能观测到theta震荡,其强度和相位都会随着记录的深度(不同层)发生变化(图1A)。
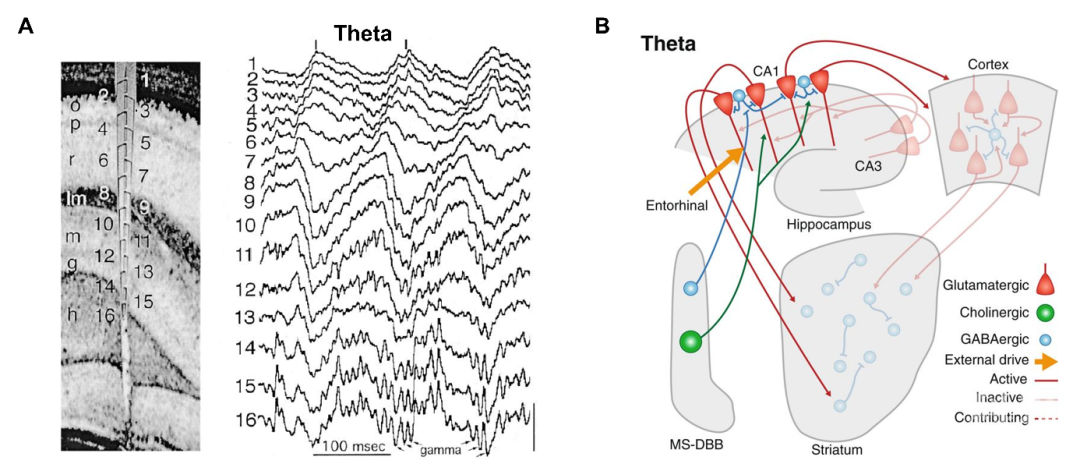
图1:大鼠内侧颞叶theta活动及其产生的机制(图1A和图1B分别引自Buzsáki,, 2002 和 Headley and Paré, 2017)
除了海马体,在海马周围的脑组织包括扣带回,杏仁核,内嗅皮层等多个区域都能记录到theta震荡。然而,这些结构都不能自发地产生theta活动。Theta活动的产生和多个皮下核团密切相关,其中内侧隔核-布罗卡斜带(MS-DBB)处的神经元控制了海马的theta震荡的产生和频率大小【2】。
MS-DBB有胆碱能神经元,γ-氨基丁酸能神经元和谷氨酸能投射神经元。用抗胆碱能药物或毒素选择性地阻断胆碱能神经元活动,能有效地降低非运动状态下或者麻醉状态下theta震荡,而运动状态和快速眼动睡眠过程中的theta震荡不受影响【3】( 图1B)。
用光遗传的手段选择性地抑制γ-氨基丁酸能神经元的活动能够降低运动相关和快速眼动睡眠中的theta活动。MS-DBB作用于海马的中间神经元,并通过周期性的抑制海马锥体细胞来控制theta节律。
Theta震荡在记忆中的作用机制
关于theta震荡在记忆中的作用机制有多种不同的理论模型,其中主要的两种: (1) Hasselmo等人提出的编码和提取相位分离模型(SPEAR model)【4】, (2) Buzsáki提出的时间编码模型【5】。SPEAR模型认为,在编码阶段的theta震荡的波峰相位时,海马CA1和CA3区域都容易接收来自内嗅皮层的新信息输入,导致这两个区域的突触后树突同时产生去极化,通过长时程促进(LTP)产生新的记忆连接。在提取阶段,尤其在theta波谷相位时,外界信息输入较弱,来自CA3的信号输入激活了CA1的活动,进而激活了与CA1连接了的大脑皮层的网络,因而提取出了之前存储的记忆(图2)。Theta处于波谷能够抑制皮层的兴奋性活动的输入,进而抑制新记忆的形成。这一模型能够解释记忆编码阶段由刺激引起的相位重置(phase-reset)。
图2:编码和提取相位分离模型 (SPEAR model)(图片引自Hasselmo et al., 2014)
时间编码模型认为theta最主要的作用是建立不同刺激在时间上的联系(图3)。这个理论是基于theta局部场电位和海马神经元放电的关系提出的。已有研究发现,在大鼠走迷宫的过程中,海马的位置细胞会依次在不同的位置放电,且这些位置细胞的发放依次锁定在theta局部场电位的不同相位上。在这一过程中,theta就好像给位置细胞的放电提供了一个桥梁,将表征不同空间位置的细胞活动连接起来,形成了一连串的空间记忆。这种记忆在时间上压缩且被反复回放。压缩后,当不同空间项目间的时间间隔足够短的时候,能够通过脉冲时间依赖的可塑性 (STDP) 来加强不同位置之间的联系,形成序列空间记忆。这一模型不仅适用于空间记忆的形成,也适用于情景记忆的形成。该模型也得到了大量的研究证据的支持,能够解释序列记忆的形成和自由回忆过程中的时间相近的词语容易被集中提取出来的现象。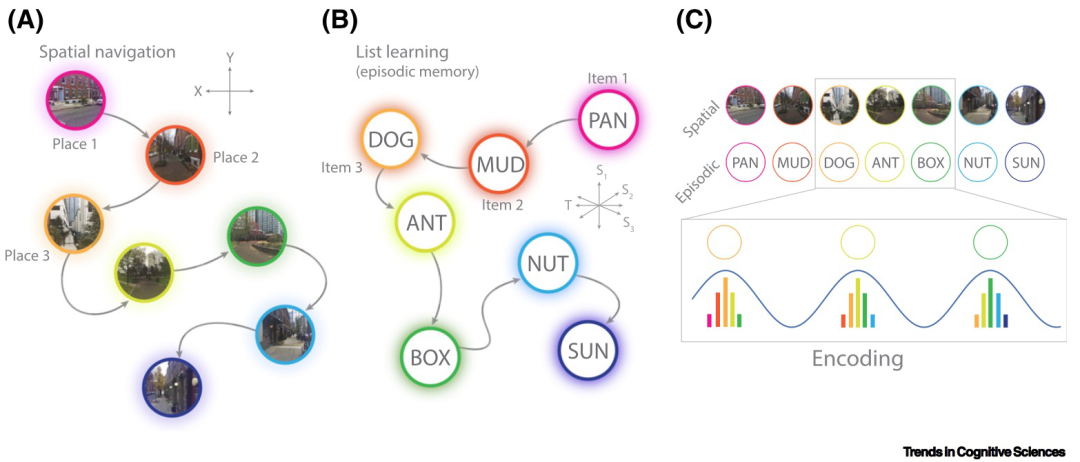
图3:通过theta活动形成联系记忆 图 (图片来自Herweg et al., 2020)
这两个模型的侧重点有所不同,第一个模型旨在说明编码和加工阶段的相位不同,更强调的是记忆网络之间的连接和theta相位的关系,没有考虑多个项目的加工过程。然而,第二个模型侧重于强调在多个项目加工过程中,如何通过theta建立项目之间的联系。这两个模型并不是互斥的。
Theta能量和记忆的关系
在过去的25年里,研究者们采用非侵入式的方法(头皮脑电和脑磁图)以及侵入式的方法(颅内脑电)来记录记忆任务(例如自由回忆,线索回忆和再认等任务)中大脑的震荡活动。主流的理论认为,情景记忆和空间导航是一样,和theta能量大小呈正相关。尽管大部分的记忆模型认为,theta效应主要在海马,但是海马和新皮层之间的投射也能驱动皮层区域的theta活动,进而通过无创性脑电被记录到。根据这一理论,内侧颞叶以及和海马交流区域其theta节律都和记忆的形成和提取呈正相关【6】。多项头皮脑电和脑磁图研究反复验证了这一观点。在记忆编码阶段,事后记住的项目(late remembered: LR)比事后忘记的项目(late forgotten: LF)有更高的theta频带的能量。在提取阶段,成功的提取或者再认的项目(Hit)比拒绝新的项目(CR)有更高的theta能量【7】(图4)。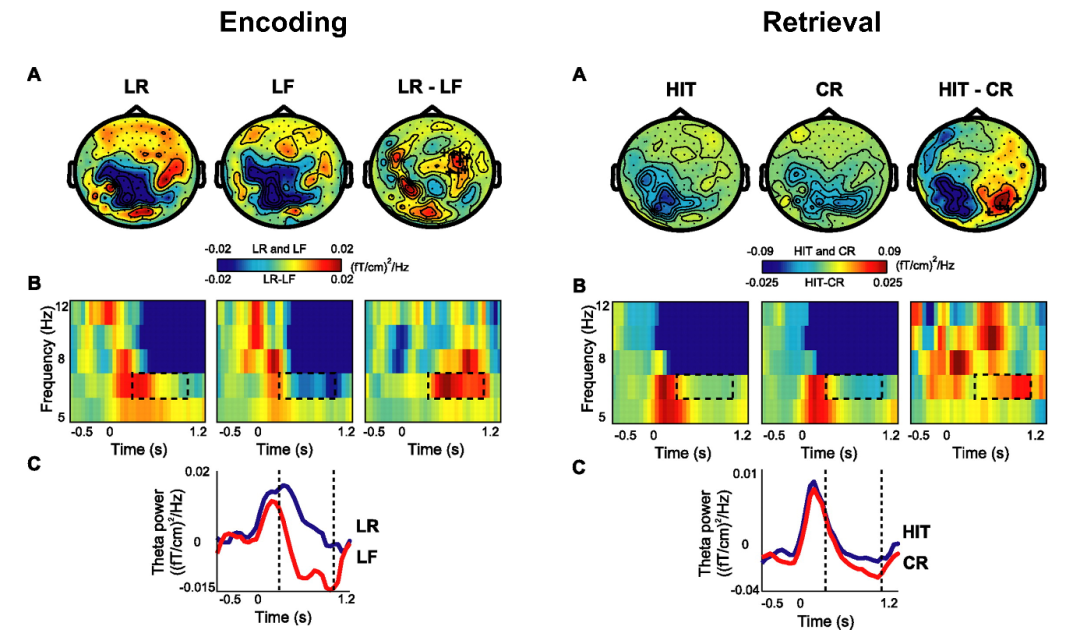
图4:编码和提取阶段记住和忘记的差异 (图片引自 Oscipova et al., 2006)
在抗药性癫痫患者上进行的颅内脑电研究发现,theta活动的模式比较复杂,有些研究发现部分电极上theta能量和记忆呈正相关,但是在另外一些电极上和记忆负相关。
和头皮脑电或者MEG发现相反,多数颅内脑电的研究报告的theta能量的大小和记忆成绩负相关,即相比是事后忘记的项目,事后记住的项目的theta能量更低。这种theta能量的降低,往往伴随着高频(30Hz 以上)能量的增加。这种现象不仅仅局限在内侧颞叶,也存在于枕叶,额叶和外侧颞叶等多个脑区【8】(图5)。
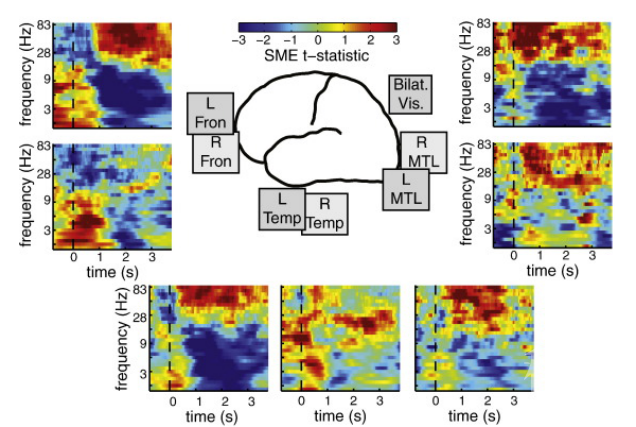
图5:记住和忘记的项目再不同频段上能量的差异 (图片引自Greenberg et al., 2015)
此外,也有少量的颅内脑电研究发现theta能量和记忆的正相关。然而这些研究不是比较记住和忘记条件下theta能量的差异,而是将theta能量的增加和记忆表现(例如,记忆的准确性,精确度等)进行关联。例如,在一项空间-物体联想记忆任务中,研究者发现相邻空间下的物体更容易一起被回忆出来,称为“空间聚类”效应。当被试成功提取相邻空间下项目的时候,被试的内侧颞叶theta能量增加。这表明,theta在提取空间线索中的作用。
关于头皮和颅内的电生理信号上观测到的theta能量和记忆关系的差异,其中一种可能的解释是头皮脑电记录到的EEG信号是局部场电位和电极间信号同步的总和。在这种情况下,即便局部场电位观测到theta能量的下降,但是由于多个电极之间信号同步行很高(正相关),在头皮检测的信号也会发现能量增加。
在灵长类动物的研究中,同步记录头皮脑电和颅内脑电发现了类似的结果。当然,目前的研究尚不能排除这些不同是否因为受试者群体不同。今后的研究可以通过同时记录癫痫病人头皮和颅内脑电的变化,来进一步排除这种可能。
Theta相位和记忆的关系
上述的讨论主要探讨theta能量和记忆的关系,关于theta相位和记忆的研究主要集中以下方面:(1)Theta 相位-相位同步能够有效的促进跨脑区的信息交流。头皮脑电研究发现,事后记住的项目比忘记的项目,其编码阶段额叶和顶枕叶之间的相位同步性增加。颅内脑电研究也发现在自由回忆和空间导航任务中,多个脑区之间的相位同步增加。(2)加工项目信息的神经元集群的高频(gamma)活动的能量和theta相位进行跨频带的耦合,能够促进空间记忆和序列项目的加工【9】,这和时间加工模型的假设是相呼应的。此外,这种theta-gamma耦合的强度能预测个体的工作记忆的成绩。未来的研究方向
本文探讨了编码和提取不同相位模型以及时间编码模型这两种theta作用于记忆的工作机制,然而这两者之间的关系并不明确。未来的研究需要通过考察并对比记忆的编码和提取,加工单个项目和多个项目,情景记忆和空间记忆过程中的活动,揭示这两种模型在记忆中的关系。Theta能量和记忆的关系在头皮脑电和颅内脑电中的差异仍有待进一步的研究。未来的研究可以通过同时记录癫痫患者头皮脑电和颅内脑电的信号,来进一步确定差异产生的原因。已有的研究大多数平均了记住和忘记条件下theta能量,然后再对比它们的差异。这种分析忽略了单个试次的theta活动,并且容易混淆一般的认知过程和记忆加工过程中theta的差异。今后的研究应该更多的关注单个试次中窄带的theta信号,分离出项目引起的theta能量的变化和由一般认知过程(注意,情绪)等引起的theta能量变化。目前,已有一些新的数据处理方法,例如MODAL【10】,能够有效的探测单个试次不同时间段的theta活动。参考文献:
1.Landfield, P. W., McGaugh, J. L. & Tusa, R. J. Theta Rhythm: A Temporal Correlate of Memory Storage Processes in the Rat. Science 175, 87–89 (1972).
2.Buzsáki, G. Theta Oscillations in the Hippocampus. Neuron 33, 325–340 (2002).
3.Headley, D. B. & Paré, D. Common oscillatory mechanisms across multiple memory systems. Npj Sci. Learn. 2, 1 (2017).
4.Hasselmo, M. E. & Stern, C. E. Theta rhythm and the encoding and retrieval of space and time. NeuroImage 85, 656–666 (2014).
5.Buzsáki, G. Theta rhythm of navigation: Link between path integration and landmark navigation, episodic and semantic memory. Hippocampus 15, 827–840 (2005).
6.Herweg, N. A., Solomon, E. A. & Kahana, M. J. Theta Oscillations in Human Memory. Trends Cogn. Sci. 24, 208–227 (2020).
7.Osipova, D. et al. Theta and Gamma Oscillations Predict Encoding and Retrieval of Declarative Memory. J. Neurosci. 26, 7523–7531 (2006).
8.Greenberg, J. A., Burke, J. F., Haque, R., Kahana, M. J. & Zaghloul, K. A. Decreases in theta and increases in high frequency activity underlie associative memory encoding. NeuroImage 114, 257–263 (2015).
9.Lisman, J. E. & Jensen, O. The Theta-Gamma Neural Code. Neuron 77, 1002–1016 (2013).
10. Watrous, A. J., Miller, J., Qasim, S. E., Fried, I. & Jacobs, J. Phase-tuned neuronal firing encodes human contextual representations for navigational goals. eLife 7, e32554 (2018).
来源:brainnews brainnews
原文链接:https://mp.weixin.qq.com/s?__biz=MzI2ODEyOTE3OQ==&mid=2649571397&idx=1&sn=91607486199fa5105a28188e14eab083&chksm=f2eda6b1c59a2fa78a88e09095cb0cad67170678e9454eed8b46e55c99ee914eea74a1f700ae#rd
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn

睡觉时提示学习过的内容有助于增强记忆

解读大脑的认知地图:海马-前额叶环路如何通过记忆“重演”来帮助空间学习和决策

“全国科普日”重点活动 |2018年浦东新区“科普小脚丫”正式启动!

【科界传声】县级青少年科技辅导员建设亟待重视

目标导向空间记忆中海马位置细胞的序列编码研究

PNAS: 记忆丧失的凶手之一居然是海马脑区的“不速之客”——糖

X射线自由电子激光在化学与能源材料科学中的应用

Nat Neuro:科学家揭示下托-海马环路如何控制对物体位置的记忆

SCIENCE:如何利用海马改善记忆力?
Nature:调控特定类型记忆存储的神经元亚群进一步被鉴定,聚焦杏仁核和海马


科界APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号