来源:BioArt
撰文 | 章台柳同源重组
(Homologous recombination,HR)是修复DNA双链
(dsDNA)断裂、单链DNA
(ssDNA)间隙和停滞或折叠复制叉的主要途径,同时有助于端粒维持,确保减数分裂过程中染色体的正确分离。异常HR是染色体重排的基础,通常与癌症、癌症易发综合症和其他遗传疾病有关。HR修复过程中,断裂处的DNA末端通过5’到3’的链切割产生3’ssDNA悬链,可被异三聚体ssDNA结合蛋白RPA覆盖。随后,Rad51替换RPA形成PSC复合物
(presynaptic complex),Rad51是DNA重组酶Rad51/RecA家族成员,在ssDNA上形成延伸的螺旋丝。这些丝状物催化链入侵,PSC中ssDNA与同源的dsDNA供体模板配对,形成一个D-loop中间体,进而进行DNA双链断裂修复、合成依赖性链退火和断裂诱导复制等过程。同源重组的关键步骤是损伤DNA必须与同源供体dsDNA配对,即同源搜索
(homology search)【1】。同源搜索的重要特征是相关搜索中间体的瞬时性和高度动态性,为全面理解同源搜索带来挑战。目前同源搜索的原理包括3D的段间转移、短距离1D的扩散及长度相关的同源性识别的组合
【2】,但这些机制只考虑了Rad51/RecA重组酶的基本特性,忽略了辅助因子的作用。所以,我们对于HR过程中DNA序列的配对过程的分子机制缺乏全面的认识。
Rad54是Swi2/Snf2家族成员,促进链入侵、催化核小体重塑和从dsDNA中移除Rad51,其缺失导致线虫、小鼠和鸡细胞对DNA损伤的敏感性增加,人的RAD54突变和乳腺癌、结肠癌、淋巴癌和其他癌症相关。现有研究显示Rad54参与同源搜索过程。首先,Rad54中dsDNA依赖性ATP水解酶活性是被Rad51的ssDNA激活的。第二,Rad54可导致dsDNA对ssDNA特异性核酸酶敏感,且这种活性可被Rad51的ssDNA刺激;这两种效应都不依赖于DNA序列同源性,表明Rad54作用发生在同源识别之前。第三,HR的早期阶段与Rad54依赖的染色体迁移率增加相一致,反映同源搜索过程中远距离DNA移动。第四,Rad54促进异源模板和PSC的联合。最后,RAD54缺失扰乱体内的同源搜索
【3】。然而,目前缺乏对Rad54在同源搜索中潜在作用机制的研究。
近日,来自哥伦比亚大学的Eric C. Greene在Cell杂志上发表文章
Rad54 Drives ATP Hydrolysis-Dependent DNA Sequence Alignment during Homologous Recombination,
利用单分子成像技术证明Rad54将同源搜索的基于扩散的途径转变为通过ATP依赖的分子运动驱动机制,主动促进DNA序列的配对过程。同时,Rad54可破坏供体模板链的配对,确保同源搜索在RPA结合的迁移性DNA气泡状结构中进行。

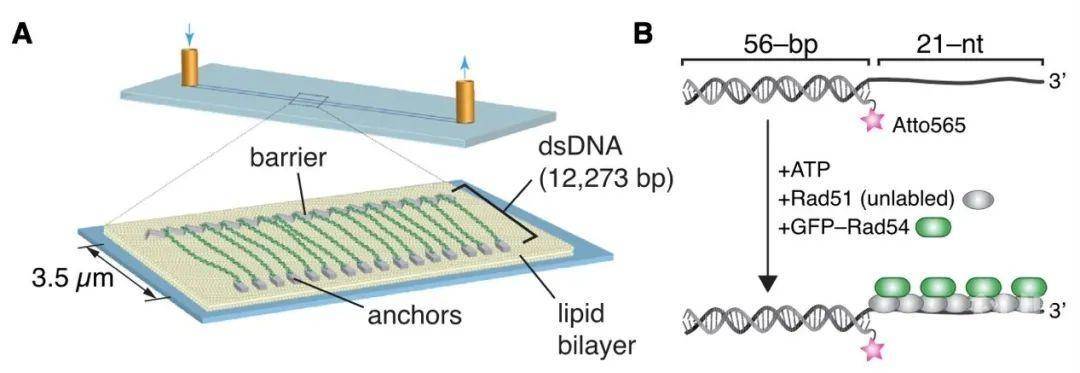
研究人员首先利用全内反射荧光显微镜
(TRIF)建立出同源搜索的可视化技术,同时构建出同源搜索反应体系:线性的CURMID
(pUC19质粒加入9.5kb的λ噬菌体DNA)一端用biotin标记、一端DIG标记,固定在样品室
(sample chamber)表面,PSCs和Atto565-DNA复合物,供体dsDNA,GFP-Rad54,RPA等。GFP-Rad54、Atto565-DNA和供体dsDNA三者共定位,且依赖于Rad51。值得注意的是,同源搜索反应中PSCs不是静止的,而是沿着供体dsDNA进行广泛的1D易位,且ATP水解酶缺陷型Rad54-K341R突变体没有易位现象,说明同源搜索需要Rad54的ATP水解酶活性。平均PSC易位速度是151 ±75bp s-1,平均持续性
(processivity)为5.2 ±1.9kb。当PSCs与更长的90-、150-、1000-nt ssDNA底物形成复合物时,其易位速度增加到302 ±172、477 ±266、592 ±421,同时Rad54的ATP水解速率并没有增加,表明更长的PSCs底物有助于Rad54更高效地ATP水解以便于供体dsDNA上的1D易位。即Rad54与PSC共定位,并促进PSC可快速地沿着供体dsDNA进行ATP依赖的易位活动。
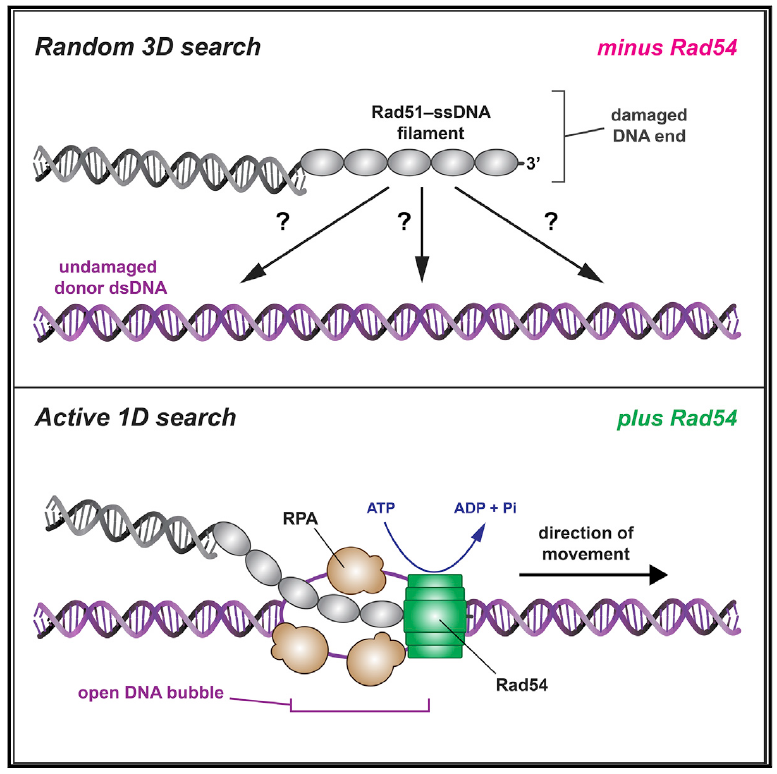
如果1D易位代表了真正的同源搜索行为,那么PSCs在供体dsDNA分子中遇到同源片段时将会中止。实验显示PSC在供体dsDNA的同源靶点区域积累。
PSCs第一次遇到同源段的识别效率与同源物的长度成比例,同源物长度越短识别效率越低,当PSCs-完全互补的21-nt ssDNA复合物,其识别效率为33%±4.0%;当21nt中只有15或9nt的同源段时,其识别效率为17.8%±7.5%、9.8%±3.0%;当21nt中没有同源段时,PSCs无法识别靶点区域,无法形成D-loop。观察发现65%的PSCs易位不发生反转,剩下的PSCs经历一次或多次反转,这提示我们如果PSCs在第一次未能识别同源段,那么它可能对供体dsDNA进行重复识别期间反转方向进行识别同源段。PSCs中ssDNA仅与供体dsDNA两条链中的一条同源互补,这种不对称性是否会导致PSCs表现出相对于同源识别的方向性偏差?研究显示用尾部双链制备的PSCs不论易位的方向,均能够有效地识别同源靶点;利用90-、150-、1000-nt ssDNA制备的PSCs可从任一方向进行同源识别,但从错误方向接近时识别的效率降低。进一步分析显示,易位速度和以错误方向接近时识别同源性的概率之间有反比关系。这表明同源搜索时,PSCs可同时扫描供体dsDNA的两条链,尽管从错误方向接近时识别同源性的能力可能会因为速度高而受损。
同源搜索反应体系中RPA对D-loop的高效形成具有重要作用。追踪RPA-GFP的动向,PSCs与RPA-GFP共定位,共同扫描供体dsDNA。82%的所有PSCs
(静止和易位)和64%的易位PSCs与RPA-GFP共定位,而Rad54-K341R突变体或添加不能水解的ATP类似物导致共定位显著减少、阻止PSCs易位和D-loop的形成,即RPA与PSCs共定位与Rad54动力活性有关。有趣的是,去除RPA并不影响PSCs易位的发生,但显著降低第一次遇到同源段的识别效率。进一步分析显示,去除RPA导致PSCs从错误方向接近同源段的识别效率大大降低,且错误方向的识别比例比正确方向降低了65%。即无论链的方向,RPA确保PSC可同时对供体dsDNA的两条链进行同源性取样。
之前的研究显示,Rad54单独也可在dsDNA上易位。研究人员利用可视化技术验证了GFP-Rad54在dsDNA的易位,且易位速度96 ±61bp s-1,平均持续性
(processivity)为5.0±3.2kb,但需要增加5倍的GFP-Rad54浓度才能观测到与dsDNA的结合,易位速度也低于PSC复合物体系。RPA与单独易位的GFP-Rad54共定位,但对其易位参数没有显著影响,两者的共定位需要Rad54的ATP水解酶活性。然而,RPA与Rad54间不存在蛋白-蛋白间互作,这与之前的报道相一致。Rad54属于Swi2/Snf2家族成员,可重塑核小体环境。利用尾部双链21nt-ssDNA和Atto565-H2A/Alexa488-H4核小体进行分析发现,PSC在遇到核小体时,要么停滞
(6%),要么重塑核小体
(51%),要么绕过核小体
(43%)。重塑事件可细分为核小体滑动
(83%,Atto565-H2A;74%,Alexa488-H4)和核小体从供体DNA中完全解离
(17%,Atto565-H2A;26%,Alexa488-H4)。即遇到核小体时,相当一部分PSC会绕过核小体,但对核小体本身没有明显影响,尽管在某些情况下,PSC在绕过核小体前会暂停相当长时间。
总的来说,
研究表明Rad54从根本上改变了同源搜索机制,从一种基于被动扩散的基本HR机制特征到一种基于ATP依赖的分子运动的主动机制,且PSC主动扫描双链DNA寻找潜在的序列同源性,为我们拓宽对同源重组的理解。来源:BioGossip BioArt
原文链接:https://mp.weixin.qq.com/s?__biz=MzA3MzQyNjY1MQ==&mid=2652491697&idx=1&sn=e8bf471f31616e1838c85404061a2f5d&chksm=84e25c05b395d5138176ab676c2d6befe0413156ac59a1929d734b37aee88da50928f328f994#rd
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn







 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号