科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2020-07-22
来源:生物通
哈佛大学著名的David Liu实验室在2016年利用Cas9融合蛋白开发出单碱基编辑器,能够大大提高碱基编辑的效率。自此,研究人员不断开发碱基编辑工具,可在动物、植物和微生物中实现点突变。
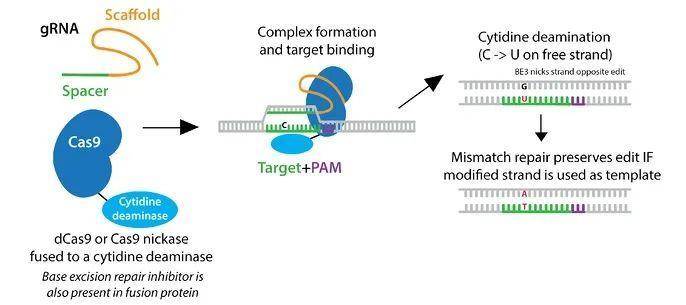
胞嘧啶碱基编辑器的示意图(图片来自Addgene)
人类的许多遗传病是由基因组中的突变引起的。因此,科学家在不断寻找工具去编辑或修复遗传代码中的这些错误。CRISPR/Cas9便成为最有希望的技术之一。利用这把“剪刀”,研究人员能够改变或删除某个基因。然而,CRISPR的应用也面临一个问题,那就是同源定向修复(HDR)的编辑效率太低。
为了应对这一问题,哈佛大学著名的David Liu实验室在2016年利用Cas9融合蛋白开发出单碱基编辑器,能够大大提高碱基编辑的效率。自此,研究人员不断开发碱基编辑工具,可在动物、植物和微生物中实现点突变。如今,碱基编辑已经广泛应用于基础研究、合成生物学和作物改良,引起了学术界和工业界的极大兴趣。
早期的碱基编辑器
David Liu实验室最初利用大鼠的胞苷脱氨酶APOBEC1和dCas9创建出第一代的碱基编辑器(BE1),能够将胞嘧啶(C)转化成尿嘧啶(U),从而实现C→T(或G→A)的碱基替换1。BE1表现出约5个核苷酸的活性窗口,从第4位到第8位。不过,在转移到细胞培养模型之后,他们发现编辑效率从44%陡降至0.8-7.7%,这可能是由于尿嘧啶DNA糖基化酶(UNG)参与了碱基切除修复,去除了DNA中的尿嘧啶。
于是,他们就开发出第二代的BE2系统,将尿嘧啶糖基化酶抑制剂(UGI)与dCas9相融合,从而将编辑效率提高了三倍,最高达到20%。对于BE1和BE2,由于DNA并非直接切割,因此插入缺失的形成非常少(< 0.1%)。
为了让碱基编辑的效率提高到50%以上,你需要将编辑复制到另一条DNA链上。为此,研究人员将dCas9变身为切口酶,以便模拟错配修复。BE3在未修饰的DNA链上产生切口,使它看起来像是新合成的。因此,细胞以含有尿嘧啶的链作为模板来修复DNA,并复制碱基编辑。
通过这种方式,BE3系统将各种靶点的编辑效率提高到30%以上,且插入缺失的频率仅为1.1%。这与Cas9介导的HDR相比是巨大的进步,因为HDR介导的编辑效率仅为0.5%,而且观察到多个插入缺失。不过,此系统也会带来脱靶效应,主要来自Cas9脱靶,而不是APOBEC1。
第三代编辑器的改进
日本神户大学Akihiko Kondo领导的团队随后进一步证实了碱基编辑的实用性2。他们将来自海鳗的胞苷脱氨酶与Cas9切口酶相融合,创建了Target-AID碱基编辑器。它的作用方式与BE3类似,但又不完全相同,其编辑窗口位于PAM上游18个碱基处的3-5个碱基。
David Liu实验室在2017年也生成了带有其他脱氨酶的BE3变体,包括AID、CDA1和APOBEC3G。他们发现,CDA1-BE3和AID-BE3能够比BE3更有效地编辑G后面的C,而APOBEC3G表现出较少的序列偏好。这些结果表明,特定的脱氨酶以及与Cas9的连接方式会影响编辑的作用和效率3。
之后,David Liu实验室的研究人员将重点又放在Cas9上。他们利用天然和改造的Cas9变体开发出五种PAM序列不同的碱基编辑器,从而扩大了碱基编辑的目标范围4。对于每种碱基编辑器,他们观察到的最低编辑效率约为50%,并确认融合蛋白保留了Cas9的PAM特性。他们还诱变了碱基编辑器的胞苷脱氨酶部分,产生了SpCas9碱基编辑器,其编辑窗口小到只有1-2个核苷酸。
为了减少与碱基编辑相关的脱靶效应,研究人员又建立了高保真版的BE3系统5,这个碱基编辑器采用了高保真的Cas9变体——HF-Cas9。随后,他们利用容易脱靶的gRNA来测试原始的BE3和HF-BE3。他们发现,HF-BE3的脱靶编辑比BE3低了37倍。为了进一步提高特异性,他们纯化了HF-BE3蛋白,并以核糖核蛋白颗粒(RNP)形式导入斑马鱼胚胎和小鼠内耳。
第四代的碱基编辑器
研究人员观察到,以BE3为基础的系统不仅仅会产生C→T,还会产生C→G或C→A。他们假设,这些副产物是由于尿嘧啶DNA糖基化酶(UNG)切除尿嘧啶而产生的。于是,David Liu实验室在BE3的基础上,融合了第二个拷贝的UNG抑制剂,以提高碱基编辑产物的纯度。自此,第四代碱基剪辑器BE4产生。与BE3相比,BE4的C→G和C→A产物减少了2.3倍3。
为了进一步降低插入缺失的产生,David Liu领导的研究团队将噬菌体蛋白Gam融合到BE4的N端。Gam蛋白与双链断裂的游离端结合,可能导致细胞死亡,而不是NHEJ修复,从而将修复细胞从编辑细胞中剔除。因此,Gam与BE4或SaBE4的融合使得插入缺失的频率降低1.5-2倍。
腺嘌呤碱基编辑器
以往的碱基编辑大多限于C→T的转换,或另一条链上G→A的转换。是否能够对其他碱基进行编辑?还是David Liu实验室,他们在2017年开发出一个腺嘌呤碱基编辑器,将腺嘌呤转化成肌苷,从而实现A→G的转换。由于没有已知的DNA腺嘌呤脱氨酶,他们便通过定向进化从RNA腺嘌呤脱氨酶TadA中产生。
经过七轮的分子进化,研究人员获得了四种腺嘌呤碱基编辑器(ABE)6。ABE7.10是活性最高的编辑器,在目标位置4-7的编辑窗口内显示出平均53%的编辑效率。ABE 6.3、7.8和7.9则显示出较宽的编辑窗口。与CBE不同,ABE不会在目标位点处出现A到其他碱基的转换。他们认为从DNA中去除肌苷的可能性比较小,从而阻止了碱基切除修复。
改进改进再改进
当然,任何碱基编辑器都不是完美的,于是研究人员开启了漫长的改造之旅。2018年,David Liu实验室在《Nature Biotechnology》杂志上发表文章,显示表达水平是碱基编辑效率的一个瓶颈。他们通过修饰核定位信号(NLS)和密码子使用以及脱氨酶组分的重构来优化胞嘧啶(BE4)和腺嘌呤(ABE7.10)碱基编辑器。此次产生的BE4max、AncBE4max和ABEmax编辑器可以在各种类型的哺乳动物细胞中纠正致病性SNP,并大大提高效率7。
在碱基编辑这一领域,中国学者也发表了大量成果。2018年,中科院遗传与发育所的研究人员在前期研究基础上利用Cas9变体(nCas9-D10A)融合人类胞嘧啶脱氨酶APOBEC3A(A3A)和尿嘧啶糖基化酶抑制剂(UGI),构成新的单碱基编辑系统A3A-PBE,成功在小麦、水稻及马铃薯等植物中实现比PBE更加高效的C→T单碱基编辑8。
不久前,华东师范大学的李大力和刘明耀课题组在《Nature Biotechnology》杂志发表了一种全新的双碱基编辑器。他们将人类胞嘧啶脱氨酶、腺嘌呤脱氨酶和Cas9切口酶融合在一起,开发了腺嘌呤和胞嘧啶的双碱基编辑器A&C-BEmax,它可以在同一等位基因的靶序列上实现C→T和A→G的高效转换9。
至于双碱基编辑器,同期《Nature Biotechnology》杂志上还发表了麻省总医院的另一项成果。J. Keith Joung 团队将腺苷脱氨酶(TadA)、来源于七鳃鳗的胞嘧啶脱氨酶(PmCDA1),分别融合到Cas9切口酶的N端和C端,开发出一种双碱基编辑工具SPACE(Synchronous Programmable Adenine and Cytosine Editor),可以同时引入A→G和C→T替代10。
来源:gh_c1fce5726992 生物通
原文链接:https://mp.weixin.qq.com/s?__biz=MjM5NzMwNjYyMg==&mid=2675536044&idx=2&sn=c1f9e49c52fdc942e5f1700cde38e9b8&chksm=bc51d93d8b26502b06fcbb5c82e093184380bc8189d80f858d848898c78d4e8a02f648f17748#rd
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn
重大进展!高彩霞/王延鹏开发出高精准胞嘧啶碱基编辑工具

ABE单碱基编辑器改造:变身CBE特异性编辑TC序列中的胞嘧啶
无水腺嘌呤盐酸盐

Nature Biotech | 杨力/陈佳/杨贝提出基因编辑领域发展新方向

NCB | 李大力课题组开发超高活性的系列胞嘧啶碱基编辑器(hyCBEs)

新型胞嘧啶碱基编辑器拓展C/G-T/A单碱基编辑适用范围研究获进展

新型胞嘧啶碱基编辑器拓展C/G-T/A单碱基编辑适用范围研究获进展

遗传所开发出高精准胞嘧啶碱基编辑技术

刘耀光院士团队开发高效、广靶向的植物胞嘧啶碱基编辑器PhiCBEs

复旦大学,中科院发表Nature文章 揭示一种全新的DNA修饰


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号