科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2019-06-13
来源:土壤观察
砷是一种广泛存在于自然环境中毒性较强的类金属元素,农田生态系统中的植物(尤其水稻)很容易吸收积累土壤环境中的砷。植物中的砷沿食物链向高等动物传递,威胁人类健康。除土壤本身的理化性质外,土壤中砷的生物转化也强烈影响砷的生物有效性。目前研究发现异化砷酸盐(As(V))呼吸性还原、细胞质As(V)还原、亚砷酸盐(As(III))氧化、As(III)甲基化和有机砷的去甲基化在土壤砷的生物地球化学过程中起重要作用。随着分析化学和分子生物学技术的进步,最新研究发现土壤生物也参与了砷糖、砷糖磷脂、碱甜菜碱、砷代草丁膦、硫代砷等有机砷的合成,其中三价一甲基砷和砷代草丁膦可作为新型抗生素,但其合成机制及生态学功能有待进一步研究。本文还详细介绍了为适应复合污染环境微生物通过自身的进化对抗生素和重金属形成的四种共选择抗性机制:共抗性,交叉抗性、共调控和生物膜感应,特别提出了土壤中砷污染与抗生素抗性相关联这一新的研究方向。最后对砷生物转化和砷与抗生素共抗机制的未来研究方向做了展望。
文/薛喜枚1朱永官1,2(1 中国科学院城市环境研究所,2 中国科学院生态环境研究所中心,城市与区域生态国家重点实验室)
类金属元素砷(As),位于元素周期表第四周期第VA主族,是一种广泛分布的有毒环境物质,已被美国环境保护署(USEPA)确定为人类的致癌物质。岩(矿)石风化、火山活动、矿山开采、冶炼、石油燃烧以及杀虫剂使用等自然或人为活动将砷释放进入大气、土壤和水体后,经物理、化学及生物等反应在地球圈层之间及其圈层内部进行循环。土壤中的砷可经地表径流或渗漏进入水体,或随着粉尘和烟雾等进入大气在空气中流通扩散,或通过植物的吸收积累进入食物链影响人体健康。
植物对砷的吸收效率主要取决于土壤砷的存在形态及迁移转化。土壤和水环境中的砷主要是无机态,在有氧环境中砷以砷酸盐[As(V)]的形式被吸附于铁铝等氧化物表面而不易移动,难以被植物吸收;在水稻土这种厌氧环境中,砷主要以亚砷酸盐[As(III)]形式存在,As(III)不易被吸附,流动性较大,很容易进入植物体内,所以与其它旱作农作物相比,水稻籽粒中积累了更多的砷[3]。除了土壤理化性质(如土壤通气性、pH、氧化还原电位、土壤胶体特性及吸附性等)外,土壤中的动物及微生物也通过对砷的吸收转化极大地影响着砷在环境中的迁移转化、形态毒性及生物有效性。土壤微生物对砷的转化作用主要体现在:对As(V)的呼吸还原和细胞质还原,产生移动性更强的As(III);对砷的氧化反应,增强砷与土壤矿物的吸附性进而影响砷的结合态和生物有效性;对砷的甲基化后挥发土壤砷而降低植物体内所积累砷的生物毒性。另外砷作为毒性较强的类金属元素与抗生素共存时将增强抗生素抗性基因(antibioticresistance genes,ARGs)传播扩散及其出现频率,某些有机砷如MAs(III)和砷代草丁膦还可作为新型抗生素。本文主要介绍土壤砷的氧化/还原、甲基化/去甲基化以及砷其它有机化过程的最新研究进展(图1),和四种微生物共选择抗性机制:共抗性,交叉抗性、共调控和生物膜感应,并提出了土壤中砷污染与抗生素抗性相耦合这一新的研究方向。
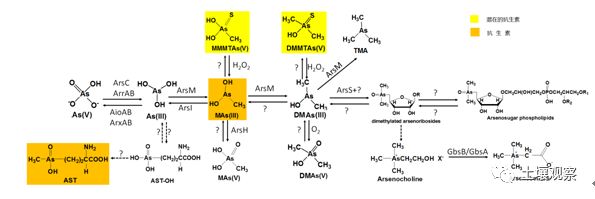
图1. 土壤中砷的生物转化途径
Figure 1. The pathway of arsenic biotransformation in soil
1. 砷的生物氧化还原
微生物砷氧化过程加速自然环境中毒性和流动性更强的As(III)向毒性和流动性更小且易吸附在矿物表面的As(V)转化,降低土壤中砷的生物有效性和毒性,因此微生物砷氧化被认为是降低砷的生物有效性,从而控制砷污染的重要途径之一。微生物的砷还原将固定于矿物中的As(V)释放并还原为移动性强的As(III),这一作用机理可运用于尾矿等区域砷污染严重的土壤砷的去除。对参与微生物砷的生物氧化还原过程的微生物类型及其氧化还原机制研究得较清楚,已有多篇综述对该过程进行了较为详细的介绍,本文不再赘述。
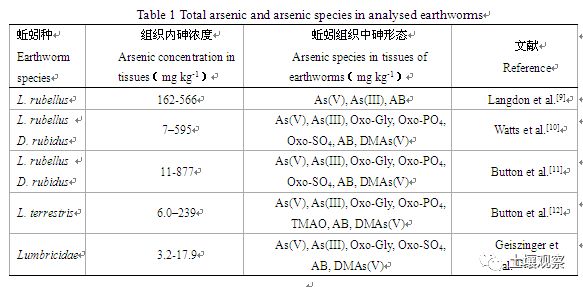
土壤动物与微生物植物一起构成复杂的土壤食物网,在陆地生态系统中扮演着重要的角色。它们在凋落物分解、土壤中元素循环、生物多样性的维持、土壤微团聚体的形成和污染物的降解等诸多土壤生态过程中发挥了关键作用。土壤动物肠道内的代谢及其营养元素富集过程改变特定元素的生物有效性。多项研究在蚯蚓Lumbricus rubellust、Dendrodrilus rubidus等的组织中检测到了As(III)、As(V)、甘油砷糖(Oxo-Gly)、磷酸砷糖(Oxo-PO4)、硫酸砷糖(Oxo-SO4)、砷甜菜碱(AB)、MAs(V)、二甲基砷酸[DMAs(V)]和三甲基砷氧化物(TMAO)等(表1),但无法确定这些不同形态砷的来源。动物体内生活着各种各样的微生物,其与宿主长期共存、协同进化,对宿主的健康、免疫、养分获取和运动等诸多方面发挥着不可替代的作用。Wang等用As(V)处理蚯蚓28天后,发现土壤中仍以As(V)为主,而蚯蚓肠道和组织中As(III)是砷的主要存在形式,砷氧化还原和解毒外排基因在所有基因中占主导,砷甲基化去甲基化相关的基因含量很低,说明在厌氧蚯蚓肠道中微生物对砷转化以氧化还原为主[8]。不论蚯蚓肠道或组织中有机砷直接来自环境还是复杂有机砷进入肠道后肠道微生物对其的降解或转化,蚯蚓等土壤动物的活动都将严重影响土壤或土壤孔隙水中砷的迁移转化、形态和生物有效性。
表 1 已分析的蚯蚓中的砷总量及形态
植物通过磷酸转运蛋白(如水稻的OsPTs)吸收环境中的As(V)。大多数植物的根细胞能将进入其内的As(V)迅速还原成As(III),例如水稻的OsACR2,A. thaliana的HAC1、Pteris vittata的PvACR2等可能参与了As(V)的还原。植物体利用水甘油通道蛋白(aquaglyceroporins,AQPs)吸收的As(III),一部分被富含巯基的多肽—植物络合素(Phytochelatin, PC)络合并被液泡膜上的ABC转动子转入到液泡内,另一部分则通过木质部往地上部运输(如水稻的Lsi2[20]和A. thaliana的肌醇转运蛋白AtINT2和AtINT4参与了该过程)。
2. 砷的生物甲基化及去甲基化
砷甲基化可以将毒性较大的无机砷转化成毒性更小的甲基化砷,虽然中间产物可能是毒性更大的MAs(III),但该中间产物极不稳定,短时间内即被氧化成MAs(V)。对厌氧微生物而言,砷甲基化过程多半产生挥发性甲基化砷从而降低环境中砷浓度,砷甲基化可能是一个解毒过程。
砷在生物体内甲基化过程是由砷甲基化酶催化,以S-腺苷甲硫氨酸(SAM)为甲基供体,巯基在还原中起重要作用。Marapakala等将真核生物Cyanidioschyzon sp. 5508的CmArsM中三个保守半胱氨酸依次突变后,蛋白荧光检测证明As(GS)3和MAs(GS)2比As(III)有更快的结合能力,这暗示着谷胱甘肽化砷可能是CmArsM的首选底物,并通过分析与苯亚砷酸盐[PhAs(III)]或还原性洛克沙砷[Rox(III)]结合的CmArsM晶体结构,推测CmArsM的砷甲基化过程是一个二硫键形成和断裂的级联反应。
土壤中存在具有砷挥发能力的细菌或真菌已被证实,真核藻类也具有将无机砷转化为气态砷的能力[27]。土壤酸碱度、氧化还原电位、含水率和温度等理化性质以及砷浓度、形态和组成等都直接或间接影响着砷的挥发。但在自然条件下土壤砷挥发量很低,可通过调节土壤环境或者通过基因工程的手段改造土壤生物,提高砷挥发水平从而达到土壤砷的生物修复。
另一方面进入土壤的大部分有机砷将被降解成毒性更强的无机砷。Yoshinaga等从MAs(V)除草剂处理的高尔夫球场土壤中分离到两株菌,一株布克氏菌(Burkholderia)可将MAs(V)还原成MAs(III),而另一株链霉菌(Streptomyces)可将MAs(III)脱甲基成As(III),这两种菌中任何一种单独存在都不能完成从MAs(V)到As(III)的脱甲基过程,暗示土壤微生物群落催化脱甲基化反应是由还原和去甲基化两个步骤组成。针对此过程的第二步,Yoshinaga等得到一个MAs(III)去甲基化酶(ArsI),该酶可以催化依赖Fe2+的MAs(III)去甲基化反应,另外,ArsI还能打断Rox(III)和其它含砷芳香族化合物的C-As键,且在原核生物中广泛存在[32]。这表明有机砷降解普遍存在于自然界中,暗示着去甲基化过程在砷的土壤循环中发挥重要作用。
3. 砷的其它有机化过程
除人工合成的有机砷外,土壤中还检测到了砷糖、砷甜菜碱及砷胆碱等多种生物来源的有机砷。近期在复杂有机砷的生物合成机制方面究取得了一系列进展。
自从1981年Edmonds & Francesconi在褐藻(Ecklonia radiate)中第一次分离鉴定到砷糖。迄今,人们已至少鉴定出了氧代砷糖(Oxo-arsenosugars),硫代砷糖(Thio-arsenosugars),三甲基砷糖(trimethylated arsenosugars)三大类共20多种砷糖。砷糖可能是砷糖磷脂合成的起点,砷胆碱和砷甜菜碱可能是砷糖降解产物。但因前期仅在基因组未测序且遗传操作复杂的海洋生物中检测到砷糖,无法开展砷糖生物合成分子机制的研究。Xue等发现纯培养的集胞藻和念珠藻均能将As(V)转化成砷糖及结构更复杂的砷糖磷脂,并基于Zhu等对砷糖合成机制的推测,,依次敲除集胞藻的arsM和arsS两个基因,用不同形态砷处理突变体及野生型,发现砷糖的合成是从DMA开始的,在砷糖合成过程中ArsM的功能是提供DMAs,ArsS可能将一个腺苷自由基转移给DMA同时将腺嘌呤切掉形成另一个自由基。双表达arsM和arsS的大肠杆菌不能将As(V)或As(III)转化成砷糖,暗示蓝细菌中砷糖的合成还需要其它所特有基因。总之,砷糖是砷生物地球化学循环的重要组成部分,它广泛存在于海洋和陆生生物中,但其存在的生物学意义和生态学功能还有待进一步研究。
砷甜菜碱是目前公认的毒性最小的含砷化合物,但它的生物学功能仍不清楚。Hoffmann等研究表明B.subtilis的甜菜碱合成酶(GbsB/GbsA)同样能将砷胆碱转化成砷甜菜碱。另外,环境中的Shewanellaputrefaciens将MAs(V)和DMAs(V)分别硫基化成毒性更强的单甲基单硫砷酸[MMMTAs(V)]和二甲基单硫砷酸[DMMTAs(V)。
4. 从砷污染到抗生素抗性基因
4.1 抗生素抗性基因及共选择抗性机制
抗生素的发现与使用拯救了千千万万的生命,是人类医疗史上划时代的成就。但随着抗生素的过度使用和滥用,ARGs的多样性与丰度在环境中不断增加。当多种ARGs同时转移到一种病原菌使其带有多重抗性,则会形成“超级细菌”,从而使抗生素不再有效。不断发展的采矿冶金、化工和农业将越来越多的重金属释放到土壤,加上抗生素的滥用,加剧了环境中抗生素及重金属协同污染的概率。重金属通过共选择的方式促进ARGs在生态系统中的扩散,并且这种共选择具有普遍性,不局限于几种重金属或者微生物。多项野外调查研究显示,土壤中的重金属与ARGs间存在明显相关性,但不同金属对ARGs丰度和多样性的影响存在差异。现在研究认为共选择抗性机制有四种(图2):
共抗性(co-resistance)是指ARGs或/和重金属抗性基因位于同一质粒、转座子、整合子上或染色体邻近位置。如Ghosh等从马流产沙门氏菌提到的质粒同时抗氨苄青霉素、砷、镉、铬和汞。然而,必须指出的是,抗金属质粒早于人类使用抗生素前已经出现。Mindlin等认为含有ARGs和汞抗性基因并携带转座子的整合子的出现是一个相对较新的现象,因为冻土源假单胞菌尽管缺乏ARGs盒但含有许多转座子与当今细菌中发现的关系密切[49]。此外,从抗生素时代前的肠杆菌科培养标本中提取的R质粒中没有可转移的ARGs[50]。这表明重金属与抗生素的协同进化一直在发生。现在需要解决的问题是早已存在的金属抗性基因是在何时如何获得ARGs的,这个时间点是否在抗生素广泛使用前,金属或类金属增加ARGs丰度和多样性的分子机制。对参与基因水平转移的可移动遗传元件大尺度测序将有助于我们深刻理解环境中金属与抗生素的共抗机制。
交叉抗性(cross-resistance)指微生物利用同一套抗性系统同时对抗生素和重金属产生抗性,当不同的抗菌剂胁迫同一种细菌时,细菌对一种抗菌剂产生耐药性的同时对另一种抗菌剂也产生抗性,即细菌使用相同机制排出结构不同的化合物。例如,Listeria monocytogenes的多药物抗性(multiple-drug resistance,MDR)泵可同时外排抗生素和重金属。编码MDR泵的基因广泛存在,但目前还没有研究能阐明这种交叉抗性的分子机制:MDR泵基因的进化、MDR泵基因的表达是否受多种抗生素或金属的调控、MDR泵是如何同时识别并排出结构差异较大的抗菌剂。

注:NOM,天然有机物 (natural organic matter);DARP, 异化砷呼吸原核生物 [Dissimilatory As(V)-respiring prokaryotes];Tetl, 四环素泵出蛋白 (tetracycline efflux protein);MATE, 多药物和有毒化合物外排蛋白 (multidrug and toxic compound extrusion); PAE1/2,马流产沙门氏菌157/158抗性质粒 (Salmonellaabortus equi157/158 resistance plasmid)[48]; Quin, 喹诺酮(quinolone);Tet, 四环素 (tetracycline); Trim,甲氧苄氨嘧啶 (trimethoprim); Chlor, 氯霉素 (chloramphenicol); AmpR, 氨苄青霉素抗性基因(Ampicillin resistance gene); Anti-As,砷抗性基因
图2 土壤微生物和动物对砷形态的转化及砷与抗生素抗性关联可能的分子机制
Figure 2. Arsenic biotransformation bysoil microorganisms and animals and proposed molecular mechanisms of coupling arsenic withantibiotic resistance
共调控(co-regulation)指一系列能对金属或抗生素暴露做出反应的转录和翻译应答系统关联在一起形成协同反应,如绿脓单胞菌的双组份传感器蛋白CzcS负责该菌对锌和亚胺培南的抗性。在大肠杆菌BaeRS调控八个基因的表达:mdtA, mdtB, mdtC, mdtD, baeS, baeR, acrD和spy,其中MdtABC与外膜孔蛋白TolC形成一个RND (resistance, nodulationand division) 型跨膜外排蛋白复合体,AcrD是AcrB的同源蛋白形成RND型转运复合体的内膜部分,Spy是一种外周质蛋白在体外表现出不依赖于ATP的伴侣活性,MdtD的功能未知。Hao等发现识别Cu(II)的MarR降低了大肠杆菌多抗生素抗性调控操纵子marRAB的表达从而导致了大肠杆菌对多种抗生素的抗性[54]。经过砷、铜和锌的诱导,Enterobacter LSJC7表现出对四环素的抗性,同时LSJC7也含有 baeRS, acrD, mdtABC, soxS, acrAB, tolC, marRAB 等基因,可能像在大肠杆菌中一样锌和铜分别通过调控BaeRS和MarR增加了LSJC7对四环素的抗性,但砷如何增强LSJC7对四环素抗性的机制有待进一步探讨。
生物膜感应(Biofilm induction)指重金属或抗生素的加入刺激微生物形成一种包裹于细胞外的多聚物,增强了其对重金属和抗生素的抗性。金属和抗生素在生物膜基质中的滞留和少量存留细胞(persister cells)的存在可能是浮游生物细胞和生物膜对高浓度抗菌剂产生时间依赖耐受性的原因[56]。向浮游生物细胞中添加金属或抗生素均可刺激胞外聚合物(extracellularpolymeric substances)的产生,从而导致细胞粘附,最终形成生物膜。另外生物膜内被保护的环境和微生物间的近距离分布也为基因的横向转移提供了便利。
Yamamura等研究抗生素对砷氧化还原和砷氧化菌多样性的影响,发现有氧As(V)还原不受抗生素影响,但有氧As(III)氧化菌对某些特定的抗生素(如氯霉素)敏感,这表明大多数抗砷菌可能同时抗抗生素。Wang等用As(V)和磺胺甲恶唑处理蚯蚓,发现砷明显增加了蚯蚓肠道微生物中特定ARGs和移动遗传元件的出现频率(数据未发表),推测进入蚯蚓肠道的As(V)被其微生物转化成毒性更强的As(III)后富集了双抗(抗砷和抗生素)的细菌,但具体机制仍需研究。总之,在土壤生态系统中土壤动物及微生物面临多重胁迫,土壤中砷的生物转化(尤其是转化成毒性更强的砷化合物)通过干扰微生物群落结构导致了ARGs丰度和多样性的改变,不同形态砷和抗生素抗性的耦合关系及机制需要进一步研究。
4.2 含砷抗生素
近期Li等提出有机砷可能是最原始的抗生素,根据Waksman对抗生素的定义,一种微生物产生的化学物质能抑制细菌和其它微生物的生长甚至杀死它们,MAs(III)可作为一种抗生素,低浓度MAs(III)即可抑制大肠杆菌的生长。土壤微生物群落中有些携带arsM的细菌产生剧毒的MAs(III)。另外已发现有四种菌P.putida KT2440, Burkholderia sp. MR1, Shewanella putrefaciens 200和Sinorhizobiummeliloti RM1021通过将MAs(V)还原成MAs(III)获得生存竞争优势。而为了在MAs(III)选择压力下生存,其它土壤细菌进化出抗MAs(III)的基因,Bacillius MD1利用ArsI将MAs(III)去甲基化成As(III)解毒,P.putida用ArsH和NADP+将MAs(III)氧化成毒性小的MAs(V),Campylobacterjejuni编码ArsP [MAs(III)排出透性酶]将细胞内的毒性强的三价有机砷排出。
砷代草丁膦(AST)是水稻根际微生物Burkholderiagladioli GSRB05产生的新型砷有机物。Nadar等发现AST抑制谷胱酰胺合成酶的活性,是一种含砷的广谱抗生素,能同时有效对抗革兰氏阳性菌和革兰氏阴性菌,其毒性与MAs(III)相当,比As(III)和草丁膦的毒性更强。P.putida KT2440的ArsN可将AST的α-氨基乙酰化阻止AST与谷胱酰胺合成酶的结合。AST具备治疗人类结核病的潜力,但该抗生素在动物和人类中的有效性和毒性还需要进一步测试[66]。总之环境中总有一些微生物产生含砷抗生素抑制或杀死其它微生物,而也有一部分微生物进化出特定的机制来适应这种环境。
5.未来研究的方向
土壤是一个极其复杂的介质,土壤理化性质影响土壤动物和微生物种类及其代谢过程,土壤中的植物、动物和微生物可以吸收转化或代谢砷。就土壤砷转化过程而言,无机砷一直是砷代谢的终点,无论是人工合成还是自然产生的复杂或简单有机砷,在自然条件下大多会被慢慢降解,最终产物是无机砷;从微生物转录调控来看,参与砷氧化还原过程的基因大多是受砷调控的,参与甲基化和其它有机化过程的基因多是组成性表达的,暗示环境中砷的氧化还原过程是砷代谢的主流。然而随着精确分析化学和现代分子生物学技术的进步,越来越多新型含砷化合物以及合成或降解这些化合物的生物或基因被发现。因这些含砷化合物在土壤中的含量较低,我们还无法知道生物为什么消耗能量产生这些复杂的有机砷、它们存在的生物学意义或生态学功能以及在食物链上的传递规律,这一系列问题有待进一步探讨。在分子水平上分析不同形态砷与土壤各组分之间的相互作用机制,土壤中微生物的种类及含量对砷化学行为的影响等有助于我们更好的理解土壤砷转化过程,为土壤砷污染修复提供重要的理论依据。另外在探明土壤微生物对砷转化分子机制的基础上进一步在群落水平上阐明其微生物生态学机制,为发展定向调控土壤中功能微生物的技术途径奠定基础。总之这些研究成果可被用来有效控制土壤-植物系统中砷的转化和积累,最终达到降低土壤砷污染风险的目标。
很明显大量抗菌药物的消耗与抗生素抗性基因的数量及种类增加显著相关,一系列其他污染物也驱动ARGs的选择。考虑到环境中普遍存在砷和抗生素等复合污染,微生物为抵抗砷和抗生素双重污染势必要进化出一套同时抗砷和抗生素的抗性系统,这些抗性菌株及其携带的ARGs更易在环境中传播扩散,将威胁人类健康和生态安全。然而现在的研究未能充分解决在生态相关框架内(如当定量ARGs在特定环境的丰度、转移和保留时区分暴露于砷的微生物种群的群落结构改变)砷暴露是增加ARGs传播和出现频率的一种重要机制。微生物基因组学、生理学和生物化学的最新进展可为精确测定砷-ARGs相互作用提供依据。此外,很少有研究将共选择的分子机制与种群和群落水平上的效应联系起来,因此有必要深入研究复合污染对环境ARGs的富集和传播机制以全面评估砷污染在维持和传播环境中ARGs的作用。并对砷和抗生素之间复杂的相互关系进行详细描述,以便系统理解ARGs的持续性和增殖。同时,一些新型的砷生物转化产物具有抗生素性能,对含砷抗生素的生物合成、毒理学及降解机制的研究无疑也是一个新的研究方向。
来源:turangguancha 土壤观察
原文链接:http://mp.weixin.qq.com/s?__biz=MzA3MDMwNTExNg==&mid=2659318673&idx=1&sn=e3168f8e9e555c17d4463482f29250d0&chksm=844b8fa2b33c06b457e363e665da1aa4e91f89e9647da53e27945645dc698b9f5aa34cc9c338&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn
微生物 调控 生物转化 甲基化 土壤结构 基因合成 土壤检测 重金属检测 重金属元素 土壤重金属污染
生物学: 咖啡害虫的致命弱点
刘沛余等-NSR:微生物如何调控生物矿化?磁小体合成的基因调控网络及模型



微生物学: 生命早期的抗生素使用可能会影响发育

【前沿追踪】空间微生物转化及应用在军民融合中的重要意义
中药牛蒡子主要活性成分微生物转化研究进展
城市环境所在城市土壤重金属污染对跳虫群落影响方面获进展

农田土壤中降低重金属污染的修复策略
Molecular Plant 出版“植物激素”专辑
昆明植物所在土壤重金属污染植物修复技术方面取得进展

华中农大在生物炭与土壤重金属污染修复研究方面取得新进展


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号