科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2020-09-17
来源:中科院之声
每年春暖花开的时节,植物园和公园常常是很多人周末出游的首选地。在那里,无论是老人还是小孩,都被五颜六色、形态各异的花儿所深深吸引。有的人在花丛之间流连忘返;有的人拿出相机、手机或画笔记录下花儿缤纷绚丽的模样;还有的人会为孩子们讲关于花的美丽故事和传说。
要说花儿为什么如此美丽,相信很多人的第一反应都是因为花瓣。的确,一般来说,花瓣是很多植物花中最显眼的器官;它们在大小、颜色、形状等方面的多样性令人惊叹。可以说,花瓣不仅是花的“调色板”,更是“造型师”。更重要的是,花瓣常以丰富的颜色和外形来吸引传粉者为植物传粉。因此,在进化过程中,花瓣的出现极大地提高了有花植物(被子植物)的繁殖效率,进而促进了它们在陆地生态系统中的壮大。所以,不同物种花瓣的色彩和形态结构是如何形成和变化的,不仅大众好奇,也是植物学家和进化生物学家关注的焦点之一。接下来,我们来讲一讲关于花瓣“造型”的故事。
被子植物中,很多类群的花瓣形态结构还算中规中矩,呈现全缘的片状结构,如常见的月季(Rosa sp.)、牡丹(Paeonia sp.)和拟南芥(Arabidopsis thaliana)的花瓣等。植物学家将这类花瓣称为简单花瓣(simple petals)。相比之下,另外很多类群的花瓣形态结构就比较有个性了,要么在边缘有裂片(lobe)、锯齿(tooth)、流苏(fringe)或其他分裂的式样,要么在其背腹面有鳞片(scale)、距(spur)、兜(pocket)或其他类型的修饰,要么以上特征兼而有之(图1)。植物学家将这类具有特殊修饰和附属物的花瓣称为复杂花瓣(elaborate petals)。其实复杂花瓣并不少见,它们在被子植物的二十多个目中都有记载,在植物-传粉者的互作中扮演着重要角色,可能促进了相关类群的辐射分化。一个典型的例子就是北美的耧斗菜属(Aquilegia)植物(图2)。在进化过程中,这类植物花瓣上距的长度变化是由传粉者的转变所介导:短距向中长距的变化是由于适应了传粉者从熊蜂(bumble bee)向蜂鸟(hummingbird)的转变;从中长距向长距的变化则是由于适应了传粉者从蜂鸟向天蛾(hawkmoth)的转变。正是花瓣上距长度的变化与传粉者之间的密切联系,使得北美的耧斗菜属植物在短时间内(约200万年)快速分化。

图1. 复杂花瓣和花瓣的复杂化方式。(A-H)具有复杂花瓣的代表物种,依次是石竹科(Caryophyllaceae)的石竹(Dianthus chinensis)、葫芦科(Cucurbitaceae)的括楼(Trichosanthes kirilowii)、兰科(Orchidaceae)的Trichosanthes kirilowii、小檗科(Berberidaceae)的小檗属一种(Berberis sp.)、毛茛科(Ranunculaceae)的华北耧斗菜(Aquilegia yabeana)、兰科的彗星兰(Angraecum sesquipedale)、毛茛科的菟葵(Eranthis stellata)和兰科的硬叶兜兰(Paphiopedilum micranthum)。(I)花瓣复杂化方式的模型。(Zhang et al., 2019.)
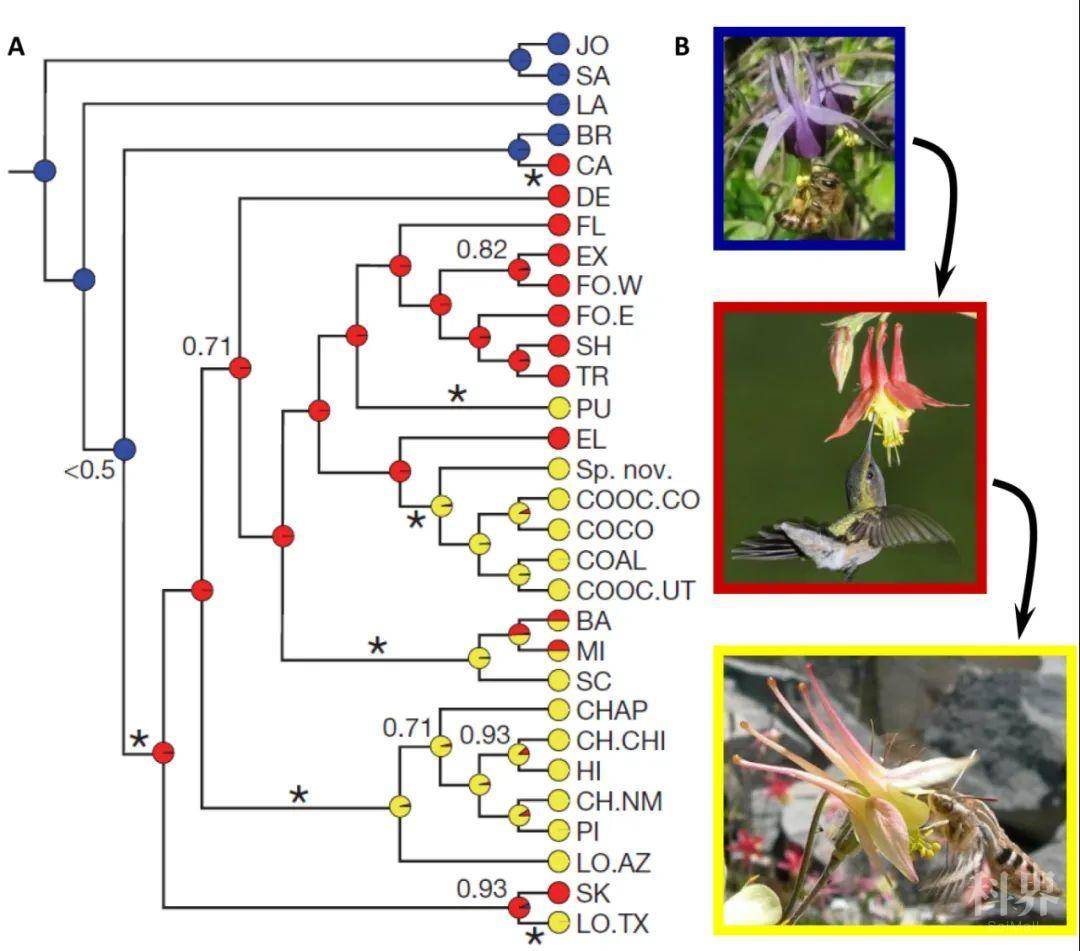
图2. 北美耧斗菜距长度的多样性与传粉者之间的关系。(A)北美耧斗菜属植物的进化关系。图中不同节点处的圆圈表示该处为熊蜂(蓝色)、蜂鸟(红色)和天蛾(黄色)传粉的概率。(B)三种具不同距长的耧斗菜及其相应的传粉者。该图改自Whittall & Hodges, 2007。
那么,形态各异的花瓣是如何形成的呢?其实,无论是简单花瓣还是复杂花瓣,它们都要经历起始(initiation)、身份决定(identity determination)、形态建成(morphogenesis)和成熟(maturation)这四个发育过程。此外,复杂花瓣还要经历一些特殊的发育过程。前人的研究发现,导致花瓣复杂化的机制主要是边缘复杂化(marginal elaboration)和腹面复杂化(ventral elaboration),还可能会涉及到特化性状的形成(图1)。例如:石竹科(Caryophyllaceae)石竹(Dianthus chinensis)和葫芦科(Cucurbitaceae)括楼(Trichosanthes kirilowii)的花瓣可以归因于边缘复杂化,边缘会形成齿状或流苏状结构;小檗科(Berberidaceae)小檗属(Berberis)的花瓣可以用腹面复杂化来解释,腹部会产生两个凸起状的结构;毛茛科大马士革黑种草(Nigella damascena)的花瓣不仅经历了边缘复杂化和腹部复杂化,使其分别产生二裂的裂片和上唇(upper lip),还形成了假蜜腺(pseudonectary)、长表皮毛(long hairs)和短表皮毛(short trichomes)等特化性状(图3)。

图3. 大马士革黑种草的复杂花瓣(左)以及与不同形态性状形成相关的代表候选基因(右)。
目前,研究人员对拟南芥这样的简单花瓣发育的分子机制已经有了较全面的认识,但是导致复杂花瓣形成的机制尚不清楚。中国科学院植物研究所孔宏智研究组长期从事花和花器官多样化的分子机制研究。近期,该团队以具有典型复杂花瓣的大马士革黑种草为研究材料,通过详细的比较转录组-表达谱及表达和功能研究,解析了花瓣复杂化的分子机制。
他们鉴定了在花瓣中特异和特异高表达的基因和程序,发现了复杂花瓣形态建成所必需的基因和程序。值得注意的是,在这些基因和程序中,除了一些是与简单花瓣所共享的(如花瓣身份决定、背腹性决定、细胞分裂和花青素合成等程序)以外,还有很多是复杂花瓣特异的(如分生组织发育、雄蕊发育、蜜腺发育和表皮毛分化等程序)。这表明后者很可能对复杂花瓣的形成具有重要作用。
通过将这些基因的表达变化与花瓣的发育过程相关联,他们筛选出30个可能控制花瓣边缘复杂化、腹面复杂化及特化性状形成的候选基因(图3)。例如,花瓣下唇的二裂可能与 CUC 类基因在花瓣中表达有关,花瓣上唇的突起可能与 KNOXI 类基因的再次激活有关,长、短表皮毛的分化可能分别由GL3和 LMI1 类基因控制,蜜腺的形成很可能由 STY 类基因决定。
为了验证这些预测是否可靠,他们对其中一个候选基因(即LMI1)进行了表达和功能验证(图4)。已知该基因在其他物种中控制表皮毛、复叶或叶卷须的发育,但它却在大马士革黑种草的花瓣中特异表达。利用病毒诱导的基因沉默(VIGS)技术将该基因沉默会使得花瓣的短表皮毛和下唇二裂消失,表明其是短表皮毛发育和下唇二裂所不可或缺的。更重要的是,基于这一结果,以及之前发现YAB5基因的异位表达对于大马士革黑种草假蜜腺的发育至关重要,他们认为对已有基因和程序的重新招募,可能是复杂花瓣形成的关键。
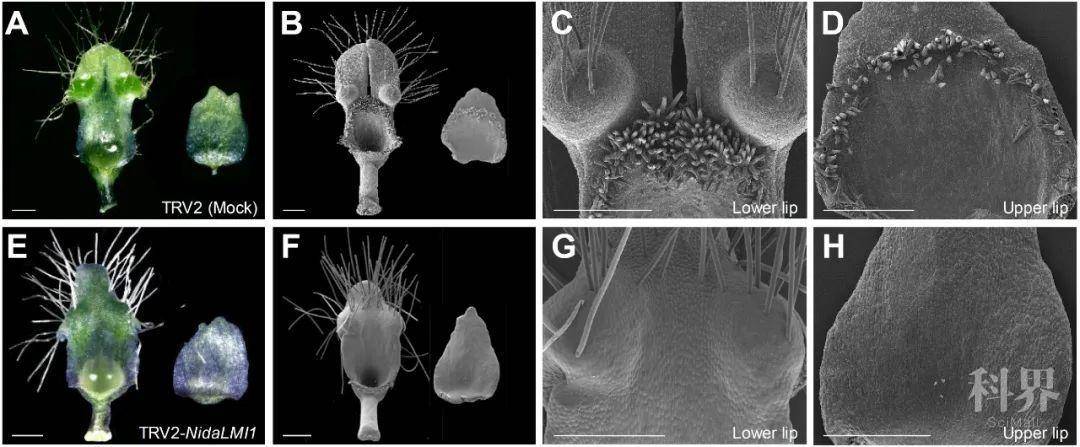
图4. 大马士革黑种草NidaLMI1基因的功能。第一行和第二行分别展示对照(Mock)和NidaLMI1基因沉默处理植株花瓣的表型。(Zhang et al., 2019.)
这项研究不仅揭示了花瓣复杂化的发育机制,澄清了复杂花瓣与简单花瓣之间的共性与特性,而且为理解其他叶性器官形态和结构复杂化和多样化的原因和机制奠定了基础。该研究成果发表于著名国际学术期刊The Plant Cell上。但目前仍有很多关于花瓣“造型”的秘密尚不清楚。例如,如果对已有基因和程序的重新招募是复杂花瓣形成的关键,那么它们是如何被招募的?黑种草属植物的花瓣呈现出非常丰富的多样性,其多样化的分子机制又是如何?相信对这些科学问题的探索和解答,会有助于人们把“花花”世界打造的更加缤纷多彩。
来源:zkyzswx 中科院之声
原文链接:http://mp.weixin.qq.com/s?__biz=MjM5NzIyNDI1Mw==&mid=2651780430&idx=1&sn=7db0b5431f55aedb95c4bd294aec9e21&chksm=bd271f1c8a50960a232e383b34e37b974771b09ad537aaf524efa17fa8a182adbbed101744d5&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn
2016年全国植物生物学大会在湖北武汉召开

Cell:番茄中法卡林二醇生物合成关键基因被鉴定,解析植物-病原菌互作新机制
第二届“分子植物”国际学术研讨会在清华大学成功召开
2015上海辰山“药食同源与植物代谢”国际学术研讨会顺利召开
《生命科学》出版植物营养与人类健康专刊

观察:“基因驱动”与合成生物学的未来

烟草糖酯合成新基因被发现

基因合成产业未老先衰?
第二届“药食同源与植物代谢”国际学术研讨会在沪举行
植物生理生态研究所科研人员发现植物抗虫调控新机制


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号