科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2020-09-23
来源:生物学霸
新型冠状病毒肺炎(COVID-19)的大流行已迅速成为全球的社会经济危机。截止目前,SAR-CoV-2 病毒在全球已感染了超 3,000 万人,并导致近 100 万人死亡。
病毒表面的刺突蛋白(S 蛋白)与宿主细胞表面的血管紧张素转化酶 2(ACE-2)结合,以启动感染。S 蛋白由两个功能亚基(S1 和 S2)组成。S1 包含受体结合结构域(RBD),让病毒初步附着到宿主细胞表面,而 S2 负责膜的融合,使得病毒进入宿主细胞。
S 蛋白不可忽视的糖基化修饰
每个 S 蛋白有 22 个 N-连接糖基化位点和至少 2 个 O-连接糖基化位点,被来源于宿主的多糖广泛糖基化[1-3]。当后代病毒颗粒形成时,病毒通过挟持宿主细胞内质网-高尔基体中间室的糖基化机制来实现糖基化。因此,S 蛋白虽然有其独特的糖基化位点,但也带有宿主细胞的糖基化模式特征。
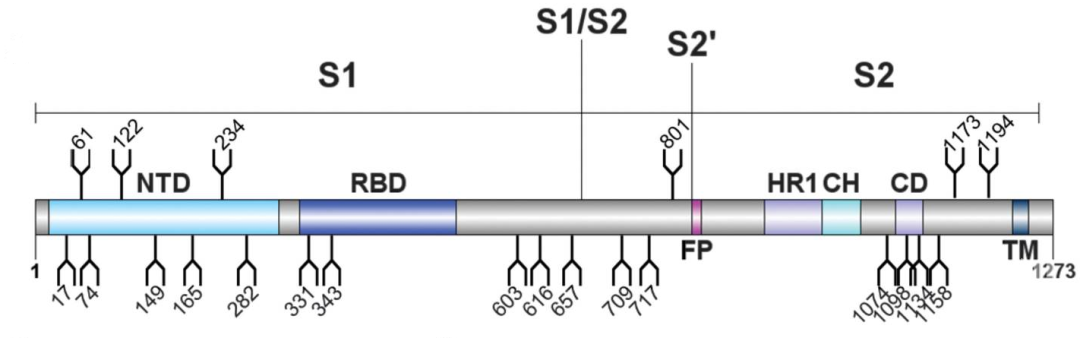
SARS-CoV-2 S 糖蛋白结构示意图,含 22 个 N-糖基化位点[2]。
S 蛋白的糖基化有助于增加蛋白的稳定性和溶解度。更重要的是,糖基化可以掩饰蛋白的免疫原性表位,从而使病毒逃避宿主免疫反应的能力得以增强。S 蛋白不仅是疫苗开发的主要靶点,也是血清学检测的重要成分。
为了促进 SARS-CoV-2 诊断和治疗的研究,R&D Systems® 的研究团队已在几种常用的宿主细胞系(HEK293 细胞、CHO 细胞和 Sf21 细胞)中表达 S 蛋白的 RBD。不同的宿主细胞具有独特的糖基化能力,因此它们表达的 RBD 蛋白有望表现出不同的糖基化特征。我们利用之前开发的聚糖直接荧光标记(DFGL)工具来分析糖基化[3],为不同目的下选择最相关的 RBD 重组蛋白提供了快速指引。
不同表达体系,RBD 糖基化特征大不同
1.O-连接糖基化
为了研究刺突 RBD 蛋白的各种聚糖结构,我们首先利用 O-聚糖特异性的唾液酸转移酶 ST3 Gal1,通过 ST3 Gal1 加入 Cy5 荧光标记的唾液酸,使得 O-聚糖可通过荧光成像仪轻松检测。
分别检测经过产气荚膜梭菌神经氨酸酶(Neu)或内切糖苷酶(EF)预处理及未经过预处理的 O-聚糖:神经氨酸酶去除糖蛋白上现有的末端唾液酸残基,并打开 O-聚糖上的位点,以便 ST3 Gal1 加入 Cy5 标记的唾液酸(Fig 1C 下)。内切糖苷酶 EF 是一种 O-糖苷酶,能够有效去除 O-聚糖简单的 Core-1 结构,但复杂的 Core-2 和 Core-3 结构不受影响(图 1C 上)。
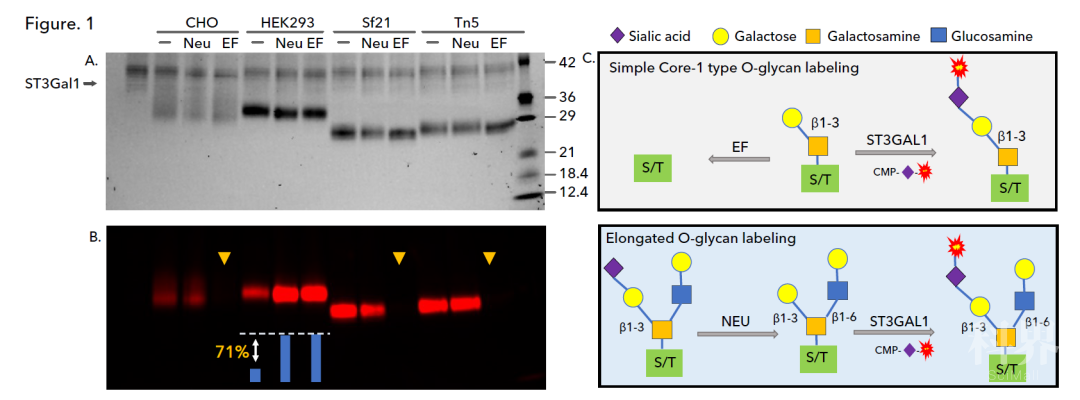
Fig 1. 利用 CMP-Cy5-唾液酸和 ST3 Gal1 检测 CHO、HEK293、Sf21 和 Tn5 细胞所表达的 SARS-CoV-2 刺突 RBD 蛋白上的 O-聚糖。通过 SDS-PAGE 分离样本,并利用 TCE 染色成像(A)和荧光成像(B)。图中显示了描述酶促反应和荧光标记的模型(C)。
首先,我们利用 ST3 Gal1 标记未唾液酸化的 O-聚糖,并检测所有四种 RBD 蛋白的荧光(Fig 1,一泳道)。神经氨酸酶的预处理显著增加了来源于 HEK293 的刺突 RBD 蛋白的标记,而来源于 CHO 和昆虫的刺突 RBD 蛋白却没有观察到变化或变化很小(Fig 1,Neu 泳道)。对来源于 HEK293 的刺突 RBD 进行光密度分析,表明荧光强度增加了 71%,这说明来源于 HEK293 细胞的蛋白中 71% 的 O-聚糖被唾液酸化。
此外,神经氨酸酶和内切糖苷酶 EF 预处理显示四种刺突 RBD 蛋白的 O-聚糖模式存在差异。主要差异在于 HEK293 刺突 RBD 上的 O-聚糖耐受 EF 处理,表明 HEK293 所表达蛋白包含其它类型的 O-聚糖(Fig 1,HEK293,EF 泳道)。CHO、Sf21 和 Tn5 所表达蛋白上的标记被 EF 完全去除(黄色箭头所示),表明这些细胞表达的 RBD 蛋白仅含有简单的 Core-1 O-聚糖。
2.N-连接糖基化
随后,我们使用 N-聚糖特异性的唾液酸转移酶 ST6 Gal1 检测了这些蛋白上的复合型 N-聚糖,同时使用 FUT8 检测了带有 GDP-AF555-岩藻糖的高甘露糖型 N-聚糖(Fig 2B)。我们将 MGAT1 加入标记反应中,以便将高甘露糖型 N-聚糖 Man5 转化成 FUT8 的底物。
结果表明,哺乳动物细胞 CHO 和 HEK293 所表达的蛋白大部分被 Cy5-唾液酸标记,表明它们主要含有复合型 N-聚糖(红色)。昆虫细胞 Sf21 和 Tn5 所表达的蛋白仅被 AF555-岩藻糖标记,表明它们仅含有高甘露糖型 N-聚糖(绿色)。
所有样本均对肽 N-糖苷酶 F(PF)处理敏感,证实了标记是 N-聚糖特异性的。有意思的是,通过光密度分析,我们发现只有 HEK293 表达的刺突 RBD 蛋白上的 N-聚糖被显著唾液酸化(33.5%),这与 O-聚糖的观察结果一致(Fig 2A)。
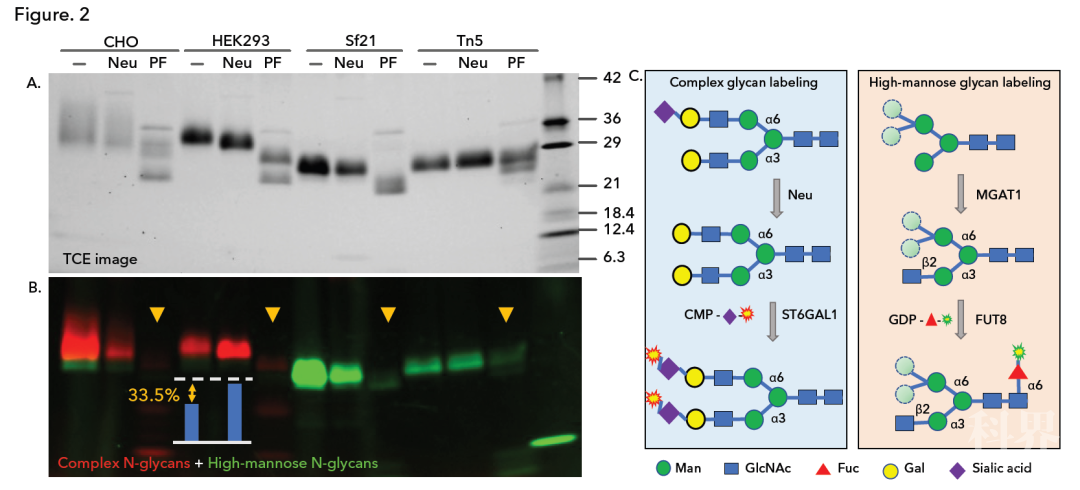
Fig 2. 利用 CMP-Cy5-唾液酸/ST6 Gal1(红色)和 GDP-AF555-岩藻糖/FUT8(绿色)分别检测 CHO、HEK293、Sf21 和 Tn5 所表达的 SARS-CoV-2 S1 RBD 上的复合型和高甘露糖型 N-聚糖。在采用产气荚膜梭菌神经氨酸酶(Neu)或肽 N-糖苷酶 F(PF)预处理之前或之后,对各种刺突 RBD 蛋白进行直接标记。通过 SDS-PAGE 分离样本,并利用 TCE 染色成像(A)和荧光成像(B)。图中显示了描述酶促反应和荧光标记的模型(C)。
通过 DFGL 检测,我们显示了四种不同细胞系所表达的 SARS-CoV-2 刺突 RBD 蛋白在糖基化上的主要差异。这些 RBD 蛋白表达的 N-聚糖和 O-聚糖类型存在明显差异(Fig 3):
01
所有蛋白都含有 O-聚糖,但只有 HEK293 细胞表达的蛋白含有 Core-1 以外的 O-聚糖,只有 HEK293 表达的蛋白存在显著水平的唾液酸化;
02
复合型 N-聚糖仅出现在 CHO 和 HEK293 表达的蛋白中。Sf21 和 Tn5 细胞表达的蛋白存在显著水平的高甘露糖型 N-聚糖,而 HEK293 和 CHO 细胞表达蛋白仅存在较低水平的高甘露糖型 N-聚糖。
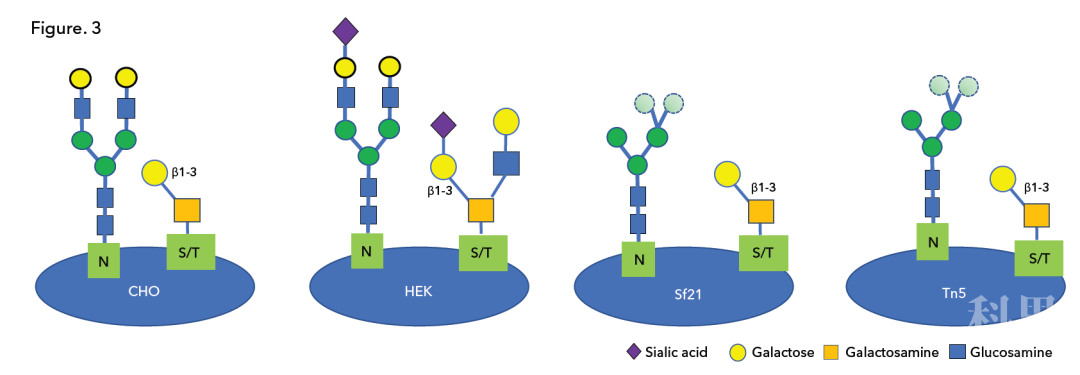
Fig 3. 四种不同宿主细胞所表达的 RBD 蛋白上的聚糖概况。CHO 表达的刺突 RBD 蛋白含有复合型 N-聚糖和 Core-1 O-聚糖。HEK293 表达的刺突 RBD 蛋白含有复合型 N-聚糖和 Core-2 O-聚糖,两者均包含末端唾液酸。Sf21 和 Tn5 表达的刺突 RBD 蛋白含有高甘露糖型 N-聚糖和简单的 Core-1 O-聚糖。
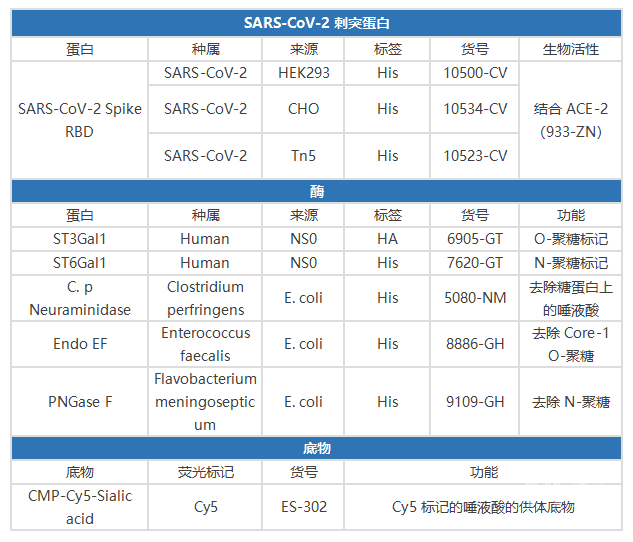
SARS-CoV-2 刺突蛋白 RBD 的糖基化研究相关工具:
参考文献:
1. Shajahan A, et al. 2020. Deducing the N- and O- glycosylation profile of the spike protein of novel coronavirus SARS-CoV-2. Glycobiology.
2. Watanabe Y, et al. 2020. Site-specific glycan analysis of the SARS-CoV-2 spike. Science.
3. Wu ZL, et al. 2019. Direct flu- orescent glycan labeling with recombinant sialyltransferases. Glycobiology, 29: 750-754.
来源:ShengWuXueBa 生物学霸
原文链接:https://mp.weixin.qq.com/s?__biz=MzA4OTQ1MTQxNg==&mid=2652088989&idx=2&sn=f4449b1f9f550c525f1c15b00944ac41&chksm=8bfdb46abc8a3d7cfe1746f7b0c4392d3224330adb174c47692a0a463ae865c951fa40f6fe7b#rd
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn
【化学生物学】氧化还原反应控制的位点特异性α2–6唾液酸酸化
自组装唾液酸糖探针实现流感病毒的“比值型”荧光检测及宿主细胞入侵阻断
王英典教授课题组闻可心、陈毅星在知名杂志上发表论文
我国科学家绘制出全球首个哺乳动物细胞图谱
天津工生所在哺乳动物细胞高通量基因编辑平台构建及碱基编辑AI预测模型方面获进展
杨辉团队证明Retron系统在哺乳动物细胞中的可适用性
Science:一“桥”飞架唾液酸,α-糖苷化变“通途”

江西师范大学孙建松课题组Angew:Pic基团指导唾液酸硫苷的α-唾液酸化反应
大连化物所实现对唾液酸糖链连接异构体的精确区分
研究提出精准可控的哺乳动物细胞基因表达系统


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号