

科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2020-11-17
植物水分生理(water relations of plant)是植物生理学的一个重要分支,研究和阐明水对植物生活的意义、植物对水的吸收、水在植物体内的运输和向大气的散失(蒸腾作用),以及植物对水分胁迫的响应与适应。水在生长着的植物体中含量最大,原生质含水量为80~90%,其中叶绿体和线粒体含50%左右;液泡中则含90%以上。
简介植物水分生理(water relations of plant)是植物生理学的一个重要分支,研究和阐明水对植物生活的意义、植物对水的吸收、水在植物体内的运输和向大气的散失(蒸腾作用),以及植物对水分胁迫的响应与适应。1
研究内容水在生长着的植物体中含量最大,原生质含水量为80~90%,其中叶绿体和线粒体含50%左右;液泡中则含90%以上。组织或器官的含水量随木质化程度增加而减少,如瓜果的肉质部分含水量可超过90%,幼嫩的叶子为80~90%,根为70~95%,树干则平均为50%,休眠芽约40%。含水最少的是成熟的种子,一般仅10~14%,或更少。代谢旺盛的器官或组织含水量都很高。原生质只有在含水量足够高时,才能进行各种生理活动。各种生化反应都须以水为介质或溶剂来进行。水是光合作用的基本原料之一,它参加各种水解反应和呼吸作用中的多种反应。植物的生长,通常靠吸水使细胞伸长或膨大。膨压降低,生长就减缓或停止。如:昙花一现,就是靠花瓣快速吸水膨大、张开;牵牛花清晨开放,日光曝晒后失水卷缩。某些植物分化出特殊的器官,水分进出造成膨压可逆地升降,使器官快速地运动。如水稻叶子在空气干旱、供水不足时,泡状细胞失水,使叶片卷成圆筒状,供水恢复后重新展开;气孔的运动是通过保卫细胞因水分情况变化而胀缩来实现的,从而调节水分散失速率,维持植株水分平衡。反之,有些器官只有失水时才能完成某些功能。如藤萝的果荚,只有干燥时才能爆裂,使其中的种子迸出;蒲公英的种子成熟、失水后才会脱离母体,随风飘荡。
水的特有的理化性质给植物带来一些好处。水的汽化热(20℃时为2 454J/g)与比热〔4.187J(g℃)〕特别高,有利于发散植株所吸收的辐射热;避免体温大幅度上升。水的表面张力、内聚力及与一些物质间的吸附力在植物体内运输中有重要意义。水能透过可见光和紫外光,使日光能透射到叶绿体上供光合作用之用,或被光敏素等吸收,引起光形态发生效应。水分子的极性造成了多种化合物的水合状态,并使原生质亲水胶体得以稳定。
陆生植物按其体内水分随环境干湿而变化的程度,可分为变水型和恒水型两大类。变水型植物如地衣、旱生苔藓和几十种维管植物(其中包括少数被子植物)。它们的细胞小而无液泡。环境干燥时,它们的含水量也随之迅速减少。大部分变水型植物在干燥时细胞均匀地皱缩,原生质凝胶化,进入隐生状态,但原生质结构不受破坏;一旦获得水分,还能迅速恢复代谢和生长。恒水型植物成熟细胞中有贮存水分的大液泡,在环境水分短暂变化时,可对细胞含水量变化起缓冲作用;有角质层阻止水分蒸发,气孔也可随时关闭;有庞大的根系和特化的输导组织以便从土壤中吸收水分输送给地上部。这些特点使恒水型植物能维持体内水分平衡,控制含水量在一定限度之内。几乎全部高等植物都是恒水型的。植物水分生理的主要内容是恒水型植物的水分代谢。1
植物细胞的水分关系植物细胞中的水分,可分为自由水和束缚水。自由水是可以移动的。生理上活跃的组织中,大部分水(包括液泡水)是自由水。束缚水是通过氢键吸附于细胞中特别是膜上的蛋白质、多糖之上的水分子,成半晶体排列,密度比液态水大。细胞壁微纤丝的纤维素和多糖胶体表面上,也有束缚水膜存在。细胞中的水分分别存在于以下几个区隔之中:①细胞壁以及细胞壁间物质,它们由亲水性的纤维素和果胶质组成。整个植株的细胞壁除根部被凯氏带隔开以外都彼此连接,形成质外体,水分可以在其中自由流动。②细胞质,其外由细胞膜(质膜)与质外体隔开,其内以液泡膜与液泡隔开。③细胞质中的各种细胞器,如核、线粒体、叶绿体等,各自有膜包围。④液泡。
各个区隔之间的膜都允许水分子通过,水透过膜的脂肪层时遇到的阻力较大。水通过的方向取决于膜两侧水势的高低。由于各种膜都是差透膜,水中溶解的离子以及糖、有机酸等有机化合物分子的透过性各不相同,因而当一个区隔中几种物质相互转化(例如大分子的淀粉或蛋白质转变为小分子的单糖、双糖、有机酸或氨基酸)时,其渗透势会发生很大变化,从而引起水的移动。植物细胞渗透势的变化可以调节水分关系。例如气孔通过调节保卫细胞钾的进出来改变渗透势,从而提高或降低膨压,使气孔开放或关闭。1
整体植物的水分代谢 陆生植物根与冠分别处于地下与地上,在通常情况下冠部向大气失去水分,根部则吸收水分,因此水的主要流向是自土壤进入根系,再经过茎到达叶、花、果实等器官,并经过它们的表面、主要是其上的气孔,散失(蒸腾)到大气中去。土壤、植物、大气形成一个连续的系统,称为土壤-植物-大气连续系。与电学中由电阻组成的线路作类比,可以把水自土壤到大气所经历的路径的各阶段看作一系列串联的阻力,如图1中的re、rb1等。通过这个途径的水的通量(F)是推动力即水势差(ΔΨ)被阻力(r)除所得的商(如式一):
陆生植物根与冠分别处于地下与地上,在通常情况下冠部向大气失去水分,根部则吸收水分,因此水的主要流向是自土壤进入根系,再经过茎到达叶、花、果实等器官,并经过它们的表面、主要是其上的气孔,散失(蒸腾)到大气中去。土壤、植物、大气形成一个连续的系统,称为土壤-植物-大气连续系。与电学中由电阻组成的线路作类比,可以把水自土壤到大气所经历的路径的各阶段看作一系列串联的阻力,如图1中的re、rb1等。通过这个途径的水的通量(F)是推动力即水势差(ΔΨ)被阻力(r)除所得的商(如式一):
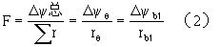
因为在稳恒条件下通过系统的各段的通量是相等的,所以可用式二表示:
即某一段的水势差(或称水势降)越大,则它的阻力也越大。从图1右侧的折线可以粗略地看出各段间的水势差,从而可以估计各段的阻力。各段阻力相差悬殊,是因为其输送过程的性质或通过的介质大不相同。水在植物体内的输送大体上可分为蒸腾、水分体内运输、根系对水分的吸收等环节。1
蒸腾作用水分通过植物表皮向大气扩散的过程称为蒸腾作用。根据扩散的通路又可分为气孔蒸腾、角质层蒸腾、皮孔蒸腾。其中气孔蒸腾在气孔开放时可占总蒸腾量的80~90%,但气孔的开张度随植株内外环境而变化。夜间或夏天中午炎热干旱时气孔关闭,阻力增加,蒸腾速率很低。1
植物体内水分运输S·黑尔斯于1727年就认识到蒸腾是引起植物体内液流上升的动力。一株植物在天气干热时,一天蒸腾失水的量可超过整个植株的重量,草本植物可超过数倍。大量蒸腾失水的补足,需有良好的通道。质外体中的细胞壁由亲水的纤维素、半纤维素和果胶质组成,其中可充满水分,又无不透水的胞膜隔断,水通过的阻力很小。在较长距离的水分运输中,木质部中的导管是主要的通道。导管由一系列中空的已死细胞组成。水以连续的水丝形式集体流动,因而阻力很小。水丝从根的中柱鞘开始,直到叶肉细胞,与整个质外体中的水形成一个连续体。高大的树木上层的叶片与根部之间的高度差可达百米。因为大气压只相当于10.34米水柱的压力,蒸腾拉力如何能将土壤中的水牵引到如此高的树冠,而树干中的水丝不断,在理论上曾经是个困难问题。英国H·H·狄克逊于1895年提出了内聚力学说,他证明:液态水的分子间有很强的内聚力。如果密闭于管道中的水柱中没有气泡或空隙,就可以象固体棒那样抵抗高达相当于300个大气压的拉力而不断。导管内有产生气泡而使水丝中断的机会,但导管细而多,组成导管的细胞或管胞之间有胞壁,从而避免了因一个小气泡出现而使整个水丝中断或气泡向上下延伸的危险。局限在少数导管内的小气泡可在夜间水分供应缓和时消失,从而使水丝恢复。
导管的壁上有各种形式的加厚结构,可抵抗蒸腾拉力产生的负压。但在蒸腾需求高时,负压仍可使导管收缩、截面积变小。树干直径每天因此表现明显的变化,可以用树径记录仪来测量并记录。因为通量与管径的四次方成正比,导管直径的收缩显著增加水的运输阻力。
白天根系吸水速率低于蒸腾失水速率时,植物体内水势下降,除导管体积缩小以外,整个质外体和与质外体接触的共质体,也都交出一些水分。有些植物果实中的水也流出,进入蒸腾流。夜间蒸腾降低后,这些部分的水量又逐渐得到补充。因而可把这些部分看作水库。但除大树和少数植物以外,库容与一天的蒸腾量相比都不大。1
根系对水分的吸收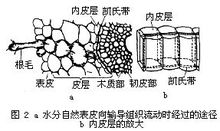 根系以其庞大的表面积,主要是根尖上着生的根毛,与土壤水分相接触。当叶片蒸腾失水时,水势比土壤水那一端的水势显著降低,在植株体内形成水势陡度,水分自土壤经根毛表面向根内流动。这种水分吸收顺着水势陡度发生,根系不起推动作用,因此称为被动吸水。当地上部不蒸腾,也没有蒸腾所造成的低水势来牵引蒸腾流时,根系也能吸收土壤中的水分,并推动水经木质部流向地上部。这种吸水过程称为主动吸水,所产生的压力称为根压。在夜间和清晨空气湿润时,某些植物(特别是它们的幼苗和叶尖)分泌出水滴,称为吐水;在切断植物的茎时,连根部分的切端上木质部处形成水珠,称为伤流。这两种现象都是根压造成的。根压一般不超过0.2MPa,远较蒸腾造成的拉力小,因此在植物吸水总量中贡献很小,也不足以将水运到离地几十米的树冠上去。主动吸水要由呼吸作用提供能量。呼吸推动离子的主动吸收,使根细胞中离子浓度高于土壤溶液之后,根细胞的水势因而下降,吸引土壤溶液中的水进入根细胞。所以这里水仍然是顺着水势差而流动的。
根系以其庞大的表面积,主要是根尖上着生的根毛,与土壤水分相接触。当叶片蒸腾失水时,水势比土壤水那一端的水势显著降低,在植株体内形成水势陡度,水分自土壤经根毛表面向根内流动。这种水分吸收顺着水势陡度发生,根系不起推动作用,因此称为被动吸水。当地上部不蒸腾,也没有蒸腾所造成的低水势来牵引蒸腾流时,根系也能吸收土壤中的水分,并推动水经木质部流向地上部。这种吸水过程称为主动吸水,所产生的压力称为根压。在夜间和清晨空气湿润时,某些植物(特别是它们的幼苗和叶尖)分泌出水滴,称为吐水;在切断植物的茎时,连根部分的切端上木质部处形成水珠,称为伤流。这两种现象都是根压造成的。根压一般不超过0.2MPa,远较蒸腾造成的拉力小,因此在植物吸水总量中贡献很小,也不足以将水运到离地几十米的树冠上去。主动吸水要由呼吸作用提供能量。呼吸推动离子的主动吸收,使根细胞中离子浓度高于土壤溶液之后,根细胞的水势因而下降,吸引土壤溶液中的水进入根细胞。所以这里水仍然是顺着水势差而流动的。
水进入根系后,主要沿着运输阻力较小的质外体运动,但根的内皮层有不透水的凯氏带将质外体的通道阻断,水只能通过属于共质体的细胞质朝木质部方向移动(图2)。穿越胞膜时经过的阻力特别大。
除根以外,植物地上部也能吸收水分。在少雨而多雾和露的特殊地区,如智利、秘鲁安第斯山脉海拔1000米高度之处的植物,水的主要来源是通过叶片吸收的。1
水分胁迫植物蒸腾失水与根部吸水之间的收支关系称为水分平衡。前者大于后者时,植物含水量下降,水势和膨压也相应降低。超过一定限度时,植物的正常生理过程就会受到干扰,甚至使植物遭受损伤,这种水分亏缺称为水分胁迫或水分逆境。土壤水分过多也对植物造成伤害,也是胁迫,但那是由于土壤渍水阻断根系的氧气供应,妨碍有氧呼吸而造成的。植物各项生理功能对水分胁迫的敏感性差别很大。生长(特别是细胞膨大阶段)最为敏感。温室中生长的,正在伸展的玉米与向日葵叶片水势只要比供水充分的叶片低0.2~0.3MPa就足以使生长明显减缓。细胞壁合成、细胞分裂、蛋白质合成和硝酸还原酶的活力等也对水分胁迫敏感。水分胁迫还引起脱落酸合成量大大增加;乙烯释放量增多;气孔关闭,光合作用减弱;以及花、果、叶脱落。中生植物萎蔫时,体内可溶性糖和氨基酸特别是脯氨酸含量明显增加。同时一些水解酶从相应的区隔中释放出来,因而产生了破坏作用。更严重的水分胁迫最终将导致生物膜系统严重损坏,造成植物死亡。
植物对水分胁迫有多种抗御的功能,就其与胁迫的关系可以分为三大类:①逃避,例如沙漠中的短命植物,在一次降雨之后,短时期(一个月)内就完成从种子萌发到开花结籽的整个周期。植物实际上不直接经受水分胁迫;②回避,植物虽经受水分胁迫,但以某些响应防止了体内不利影响的发生。例如干旱时气孔关闭,防止了水分的散失和体内水势的下降;根冠比增高使供应单位叶面积的根吸收表面积增加,从而改变供求比等;③忍耐,变水型旱生植物能忍受强度脱水,直到气干状态仍不死亡;再获雨水时能很快恢复生命活动,也称为复苏植物。恒水植物中北美南部沙漠区的Larrea tridentata,旱季中老叶和小枝脱落,只留下长成的叶和芽,含水量降到干重的50%也不引起严重损伤,雨后仍能重新生长。1
研究意义地球表面上降水量和蒸腾需求分布极不均匀,不同地区干燥度(蒸腾需求与降水量之比)相差悬殊,植物种的水分生理特点和抗水分胁迫能力,对决定其地理分布和生产力起重要作用。在农业上主要关心的是植物的产量和品质。灌溉水充裕时,追求以合理灌水来使产量最高;降水不足、灌溉有限时,则单位水量所能给出的产量最高、即水的经济利用效率最高更为重要。这在旱作农业中制定各项农业措施(包括品种的选择)中,是主要的目标。1
测定仪器茎流计茎流计应用茎热平衡原理,在不损伤植物组织的基础上,测量植物体内的液体流动,用于研究植物对水分的利用。茎流计的探头适合于直径2mm-125mm的植物茎。
茎流计探头还可配置在澳作自动气象站上与气象数据同时监测。
TDP茎流探针TDP是一种耗散式热电偶探针,用于测量较粗植物茎的茎流。将探针插入树木的韧皮部,通过测定两探针间的温度差异计算茎流。可连续记录茎流速度和液流带走的绝对热量。 TDP茎流探针还可配置在ICT自动气象站上与气象数据同时监测。
动态气孔计AP4动态气孔计能够在野外快速测量叶片的气孔导度和气孔阻力。通过气体循环,将叶夹中相对温度变化数据与特制的标定板测量的数据比较,配合温度变化数据,精确计算出气孔导度或气孔阻力。
动态气孔计不依赖于相对湿度测量的绝对精度,加之其独特的标定板技术可实时标定仪器,故野外应用时较稳态气孔计准确。
植物水势仪—压力室测量整片叶或枝条的水势。仪器将叶片或枝条夹如样品室,通过气体加压,观察第一滴组织液渗出时的压力。本公司有两种压力室: SEC压力室:可在野外和实验室应用,更适合于实验室使用,指针表读数。 Skye压力室:可在野外和实验室应用,更适合于野外使用,数字表读数。1
本词条内容贡献者为:
胡芳碧 - 副教授 - 西南大学
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号