科技工作者之家
科界APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2019-11-11
来源:BioArt植物
植物所有的地上组织都是由茎尖分生组织(shoot apical meristem, SAM)所产生的,SAM的核心成分是位于中央区域的干细胞(stem cell)。干细胞产生与维持由促进因子WUSCHEL(WUS)和抑制因子CLAVATA(CLV)组成的负反馈循环来调控【1】。促进因子WUS蛋白产生于SAM内部的组织中心(organizing center),通过胞间连丝转移到干细胞区域。抑制因子CLV3可以抑制WUS的活性,避免干细胞过量增殖,并且CLV3蛋白特异的由干细胞产生,可作为干细胞的分子标记【2】。干细胞被瞬时增殖细胞(transient amplifying cells)包围着,这些细胞可以响应生长素信号,起始器官分化【3】。在SAM,细胞内生长素的浓度是由生长素极性运输载体PIN1(PINFORMED1)调控【4】,继而生长素被核受体TIR1(TRANSPORT INHIBITOR RESPONSE1)感知,结合生长素而活化的TIR1促进生长素信号抑制因子AUX/IAA的泛素化降解,间接促进生长素响应因子ARF(AUXIN RESPONSE FACTOR)的表达【5】。SAM一方面需要持续增殖分化不断产生新的地上器官,同时干细胞又需要不断自我更新,保证在整个生命周期中维持一定数量的多能性干细胞。但关于植物SAM维持干细胞的自我更新与不断分化间的平衡的具体机制,目前还知之不多。
近日,来自于德国海德堡大学的Jan U. Lohmann团队发现干细胞促进因子WUS如同可变电阻器一般调控着生长素的信号和响应途径,从而维持SAM内干细胞的命运。相关结果以WUSCHEL acts as an auxin response rheostat to maintain apical stem cells in Arabidopsis为题发表于杂志Nature Communications上。

研究者发现生长素输入信号R2D2普遍存在于SAM,包括中心干细胞区域,但生长素输出信号DR5v2,强烈表达于分化的细胞区域,而在干细胞区域表达较弱,存在生长素信号响应极小值区。如果阻断WUS转运促使干细胞增殖分化后,中央区域的DR5v2信号明显增强。而完全阻断生长素的下游输出信号会使SAM终止,说明中央区域的生长素响应信号虽然微弱,但对于维持干细胞的命运是不可或缺的。增强生长素输出信号通常会促进器官增殖,但特异的在干细胞区域提高生长素输出信号对SAM并没有明显影响。上述结果说明干细胞命运的维持需要较低的生长素信号,但干细胞区域对较强的生长素信号响应并不敏感。
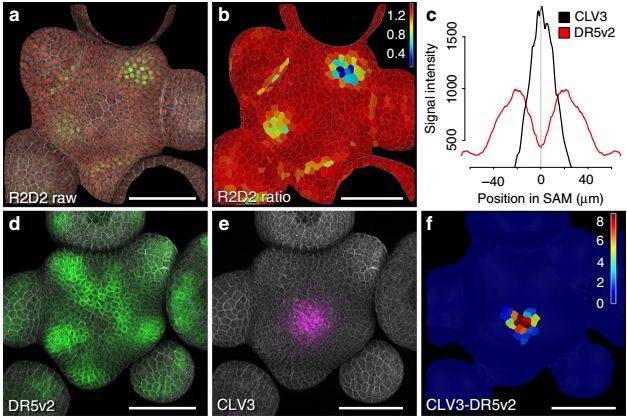
图1:茎尖干细胞存在生长素输出信号极小值区域
阻断生长素下游输出信号产生SAM终止的表型与wus突变体表型非常类似,研究者继而通过增强或削弱WUS的蛋白活性,证明了中央干细胞生长素响应极小值区域的建立依赖于WUS活性。为了进一步回答WUS是如何调控生长素的信号输出,研究者利用chip-seq实验鉴定了6740个WUS的调控位点。在WUS的靶基因中与生长素信号相关的基因高度富集,包括生长素的运输、感知、信号传导、信号响应等等方面,并且在SAM区域,WUS主要抑制这些基因的表达。WUS可以和转录共抑制子TPL相互作用,TPL进一步招募组蛋白去乙酰化酶HDACs(HISTONE DEACETYLASES),通过HDACs去除组蛋白尾巴上的乙酰化修饰来抑制基因的表达。WUS过表达株系中,有近一半WUS靶位点的组蛋白的乙酰化修饰水平降低,但大部分位点的乙酰化修饰水平降低幅度有限,说明WUS只是下调而非完全关闭靶基因的表达。利用组蛋白去乙酰化酶抑制剂TSA(Trichostatin A)的处理,进一步说明了WUS发挥转录抑制的功能依赖于组蛋白去乙酰化酶的活性。相比于野生型,wus突变体由于干细胞区域生长素响应信号的激活,SAM终止的表型对于外源添加的生长素更加敏感。
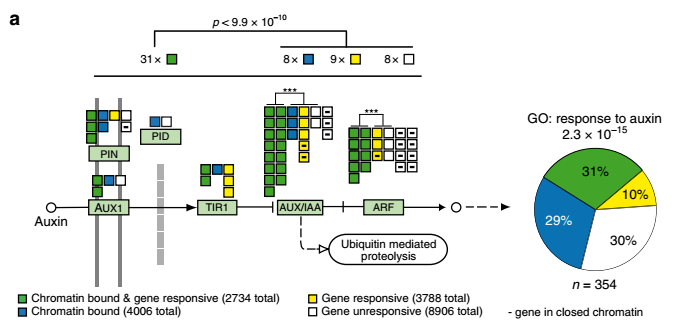
图2:WUS对于生长素信号途径的转录调控
综上所述,本研究发现由于WUS的在SAM干细胞区域对于生长素途径基因的转录抑制活性,使得该区域的生长素信号维持在较低的基本水平。一方面避免了在较高生长素信号诱导下干细胞的完全分化,另一方面基础水平的生长素信号对于维护干细胞命运,保证特定干细胞数量又是必不可少的。该研究揭示了WUS调控生长素信号途径来保持干细胞在细胞分化与自我更新间平衡的机制。
参考文献:
[1] Schoof, H. et al. The stem cell population of Arabidopsis shoot meristems in maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell 100, 635–644 (2000).
[2] Brand, U., Fletcher, J. C., Hobe, M., Meyerowitz, E. M. & Simon, R. Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science 289, 617–619 (2000).
[3] Benjamins, R. & Scheres, B. Auxin: the looping star in plant development. Annu. Rev. Plant Biol. 59, 443–465 (2008).
[4] Vernoux, T., Kronenberger, J., Grandjean, O., Laufs, P. & Traas, J. PINFORMED 1 regulates cell fate at the periphery of the shoot apical meristem. Development 127, 5157–5165 (2000).
[5] Kepinski, S. & Leyser, O. The Arabidopsis F-box protein TIR1 is an auxin receptor. Nature 435, 446–451 (2005).
论文原文链接:
https://www.nature.com/articles/s41467-019-13074-9
来源:bioartplants BioArt植物
原文链接:http://mp.weixin.qq.com/s?__biz=MzU3ODY3MDM0NA==&mid=2247492915&idx=1&sn=e45636fcdc10a15273e9438698f55cc8&chksm=fd737f54ca04f642068df45505aece5389b08ac85f9f094d2a538a9e93b66bc72c0271a903e8&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn
生长素转运蛋白PIN进化揭示生长素推动的开花植物起源进化
中国首个干细胞通用标准发布,规范国内行业发展
生长素响应的转录抑制调控网络

生长素通过细胞骨架和细胞形态调节拟南芥胚中的细胞分裂方向

3D迷你大脑,助力脑功能修复研究

Nature Plants:铵态氮介导生长素运输调节拟南芥侧根发育

中国科学家首次破译细胞“返老还童”二维码
拟南芥ARF与AuxRE重复序列的高亲和力结合调控生长素响应基因表达
皮肤变肌肉不再是幻想,科学家们做到了!
中国科协发布2016年度“中国生命科学领域十大进展”


科界APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号