科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2019-12-09
来源:BioArt
作者丨Patrick Cramer
编译、整理 | 奚望
责编 | 兮
基因表达调控决定了细胞的特征和功能。在一个物种发育的各个阶段,都有不同的基因被激活或抑制,形成不同类别的细胞。而基因的表达需要在转录水平上被精确地调控。因此转录调控机理一直是分子生物学研究的中心之一。转录过程基于RNA聚合酶。RNA聚合酶通过识别启动子序列,打开DNA双螺旋起始转录。RNA聚合酶总共有三种:1)Pol I,合成大核糖体RNA(rRNA)的前体;2)Pol II,合成信使RNA(mRNA)和大量非编码RNA;3)Pol III,合成转运RNA(tRNA)和小核糖体RNA。他们各有不同的协同因子和作用机理。
2019年8月19日,德国Max Planck研究所Patrick Cramer在Nature杂志上发表了综述文章Organization and regulation of gene transcription,主要介绍了当下对于真核生物基因调控的观点,聚焦在转录的起始(initation)和延伸(elongation)过程。作者还强调了近期关于Pol II在细胞核中与其他因子动态聚合的研究,并提出了一个模型,来解释Pol II是如何依靠磷酸化修饰在起始和延伸中在不同的因子中聚合穿梭。

启动子和增强子
转录的第一步是由RNA聚合酶结合基因起始位点的启动子。正常的染色质结构和核小体结构会阻止这种蛋白因子同DNA的接触,但是活跃启动子区域通常脱离了核小体,因此呈开放构象。Pol II一般可以识别两种启动子:一种是含CpG岛序列的启动子,通常出现在housekeeping基因的周围;另一种是含TATA盒的启动子,通常出现在细胞特异性表达的基因周围。在某一种特定细胞中,只有一部分启动子会呈活跃状态,这是由于一些特定的转录因子通过识别特定序列的方式结合在了这些启动子之上。而转录因子还可以同增强子结合,从而利用离启动子很远的增强子序列调控转录过程。
启动子识别
启动子是序列保守的DNA片段。事实上,RNA聚合酶本身并不能识别启动子序列,是其他的转录起始因子作为桥将RNA聚合酶和启动子连接起来。RNA聚合酶同这些因子形成起始前复合物(pre-initationation complex,PIC)。三种RNA聚合酶的PIC具有相似的结构。
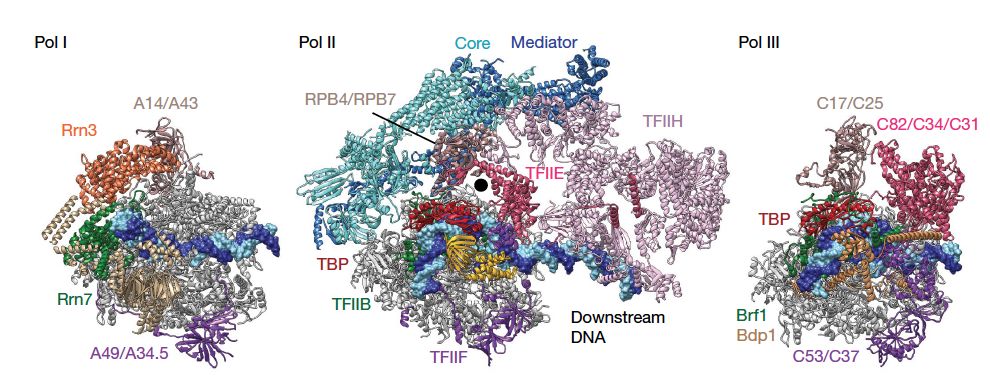
Pol II PIC主要包含第二类起始因子。其中TBP可以识别TATA box,然后结合TFIIB,接着招募RNA聚合酶以及TFIIF。TFIIB就是起到类似桥作用的因子,它还可以激活RNA的合成。然而很多的启动子缺乏TATA box这样明显的可识别序列,因此也有人认为是起始因子中的TFIID识别+1位置的核小体。启动子的识别也可能同周围环境的DNA物理性质有关,例如DNA的可弯曲性等等,这也可以解释为什么PIC组成在不同启动子上有所差别。
启动子打开
PIC的一个关键功能是将DNA双链打开。在这里,不同的RNA聚合酶有不同的机制。Pol I和Pol III将DNA识别和打开两个过程同时进行,而Pol II则需要额外的DNA转位酶XPB的帮助。XPB通过水解DNA产生能量,将DNA推到Pol III的中心。因此Pol III的转录起始多了一层额外的调控机制。
起始调控
PIC的形成在每一种RNA聚合酶系统中都受到调控。酵母Pol I的PIC需要Rrn3参与才能形成,而Rrn3的磷酸化则会阻止它同PIC结合。相应的,人类TIF-IA也起到相似的作用。Pol II PIC的形成则受到中介复合物(Mediator)的调控。Mediator有保守的头和尾结构域,它与Pol II和其他起始因子结合,尾部还可以同活跃的转录因子结合。Mediator可以激活TFIIH的CDK7结构域,从而磷酸化Pol II的CTD结构域,促使延伸过程顺利进行。
延伸调控
当RNA合成到特定长度的时候,延伸复合物就会形成。延伸复合物中的RNA聚合酶通常包含着一圈DNA-RNA杂交双螺旋。RNA每一步的合成都需要经过关闭活跃位点-形成磷酸二酯键-移动到模版链的下一位点。特定的DNA序列会导致转录的暂停(pausing),使得聚合酶反向移动或者转录终止。Pol ll常常会在核小体前发生暂停,而TFllS可以将其解救出来。
Pol II的暂停步骤也是受到精确调控的。暂停一般发生在转录起始位点下游50bp的位置上。DSIF和NELF可以稳定暂停过程中的Pol II,而从暂停中释放需要激酶CDK9(P-TEFb的一个因子)的作用。P-TEFb将DSIF,NELF和CTD都磷酸化,从而释放出一个活跃的转录复合物。磷酸化的CTD会接着募集很多其他因子形成延伸复合物,并对RNA剪切,组蛋白修饰和染色质重构都产生影响。
启动子附近的暂停会对RNA转录本产生的数量起到限制。而转录因子在起始和延伸两个阶段都能起到作用。例如,原癌基因MYC就可以促进Pol II从暂停中释放。
转录聚合
以上关于转录起始和延伸的讨论涉及到了大量的调控因子。一个自然的问题就是,这些因子是如何快速地出现在一起,并将起始和延伸的过程区分开来的呢?显微镜研究标明,Pol II的转录常常发生在细胞核内的一些“节点”中,而这些转录的焦点可能是由于相分离(phase separation)机制形成的。Pol II和Mediator在转录位点形成聚合物,Mediator和BRD4在增强子簇附近形成聚合物,而转录因子可以用它们的反式激活结构域将这些聚合物吸引到一起。作者将它们成为“启动子聚合物”,因为聚合使得转录相关因子在空间上的集中和运送变得更容易。
CDK7促发的CTD的磷酸化会使CTD脱离由相分离形成的聚合物。但是磷酸化的CTD却可以和P-TEFb形成另一种液滴状相分离复合物,从而进入到延伸聚合物中。作者将这种含有磷酸化Pol II的聚合物成为“基因体聚合物”。
转录组织的模型
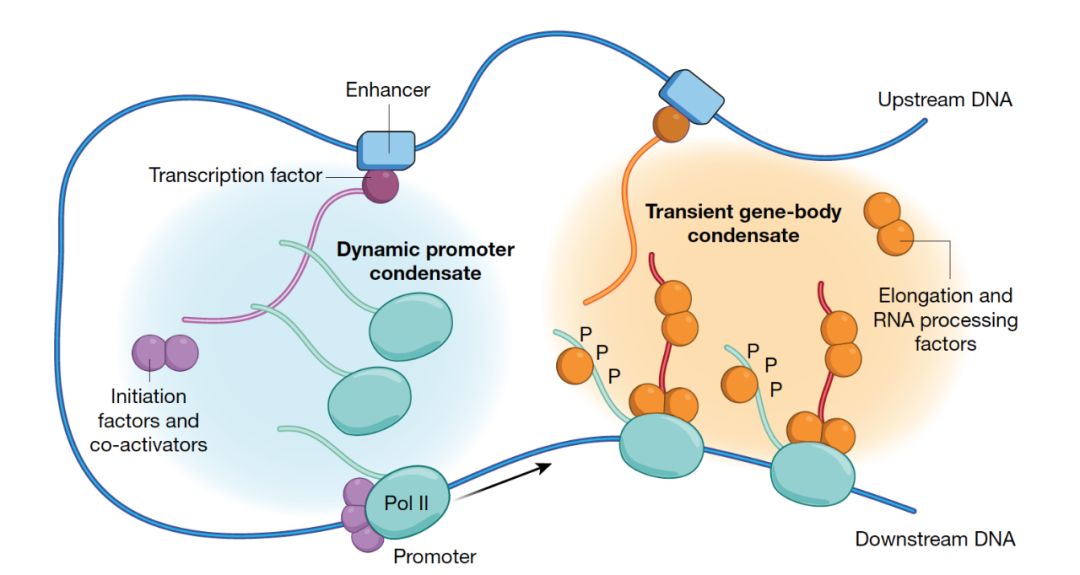
作者据此提出了一种模型,即转录因子,激活的起始因子,协同因子和未磷酸化的Pol II形成启动子复合物。而磷酸化的Pol II,新生RNA,衍生因子,RNA处理因子和协同因子则形成暂时性的基因体聚合物。起始聚合物使转录起始快速发生,而延伸聚合物则负责RNA延伸和修饰。这种机制保证了起始和延伸过程的分离。这个模型可以帮助解释细胞是如何在分化中迅速地改变基因表达,因为起始聚合物是动态的,并且依赖于细胞特异性的转录因子。一个起始聚合物也可以服务于多个活跃的基因,所以一个启动子可以影响多个基因的表达。作者认为,聚合物模型(相分离)可以成为接下来对转录分子生物学研究的框架模型。
原文链接:
https://doi.org/10.1038/s41586-019-1517-4
来源:BioGossip BioArt
原文链接:http://mp.weixin.qq.com/s?__biz=MzA3MzQyNjY1MQ==&mid=2652478646&idx=1&sn=b32e4b4c59c7d051743db57d13eb1d6f&chksm=84e20f02b39586140d16f492f34e3b0c23a35a359fe2a44f7c41cc92f81e3a9249ef608dfdde&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn

大型衰老相关基因表达谱数据库建成
中国科学院揭示热纤梭菌转录调控因子σI的独特启动子识别机制
功能相关基因协调表达的多种策略:双向启动子、基因簇、DNA 环化、操纵子及基因编辑及
通过目标启动子基因编辑及病原菌群体监控实现水稻白叶枯病持久广谱抗性

PBJ:四川大学开发出高效的双向绿色组织特异型启动子
Nat Comm :科学家捕获代谢适应过程增强子启动子网络变化

夜班影响:基因表达不能适应新的睡眠模式
Nat Microbiol:新研究揭示双向启动子在生命三界中普遍存在

全面分析拟南芥、玉米和高粱核心启动子,助力设计新启动子和作物改良
“常绿”基因OsSGR启动子的优良变异延缓叶片衰老并提高水稻产量


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号