科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2020-06-22
来源:BioArt植物
染色体浓缩是细胞分裂的前提基础,有丝分裂和减数分裂都需要将复制后的细长染色体浓缩成棒状的染色体,进而保证后期的正确分离。相比之下,有丝分裂中染色体从前期到中期是快速的一步浓缩,而减数分裂包括减数分裂I和II,其中减数分裂I是该细胞分裂方式所特有,前期I又包括细线期、偶线期、粗线期、双线期和终变期,过程主要涉及到父母同源染色体之间的配对、联会和重组。为了保证这些核心事件的有序进行,染色体的浓缩是逐步进行。研究表明染色体浓缩主要由浓缩和粘黏蛋白复合体控制,但真核生物减数分裂进程中如何调控浓缩蛋白复合体,从而完成染色体的逐步浓缩机制不清楚。王应祥和马红团队2016年揭示了植物减数分裂细胞特异编码PHD结构域蛋白MMD1,其可能通过PHD识别组蛋白H3K4me3直接调控浓缩亚基CAP-D3的表达,从而控制了染色体的前期浓缩进程(Wang et al., Plant Cell)。但是,MMD1调控CAP-D3的具体分子机制不清楚。
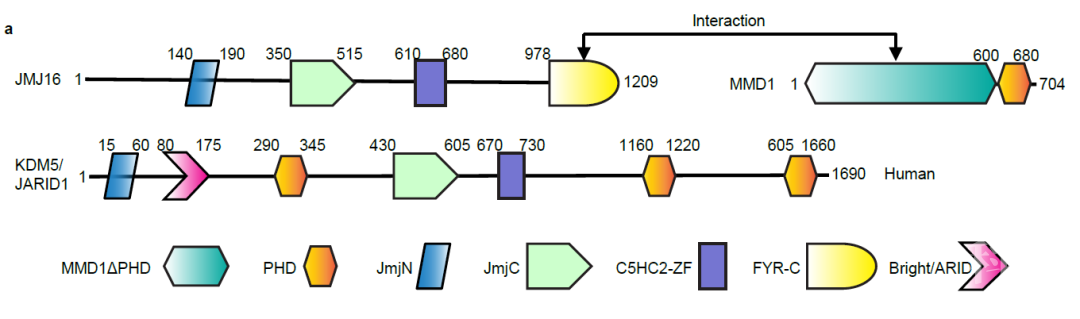 图1 MMD1和JMJ16蛋白的结构域及相互作用区域
图1 MMD1和JMJ16蛋白的结构域及相互作用区域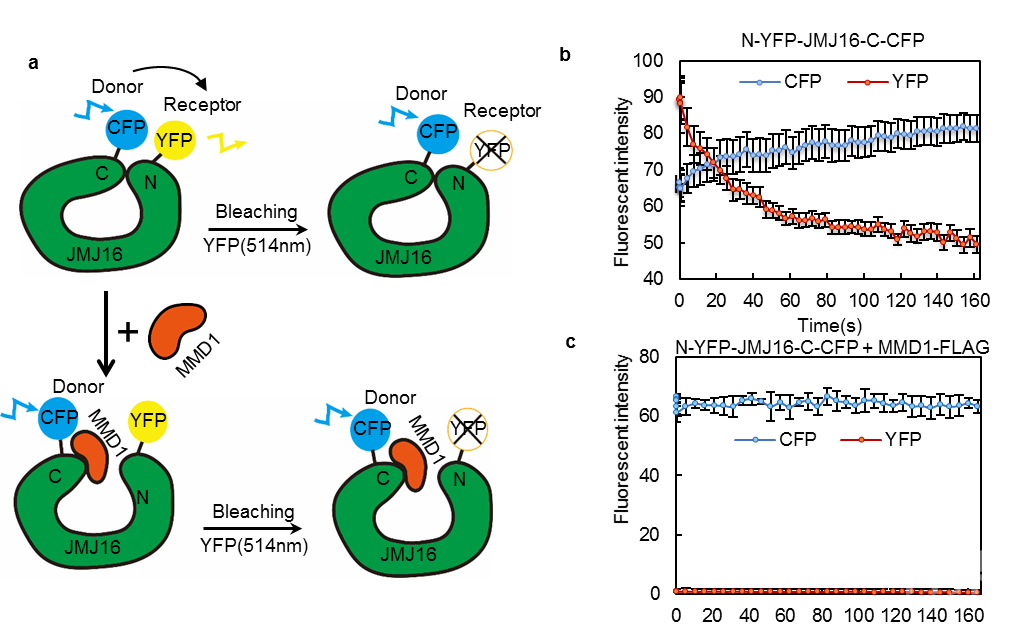 图2 JMJ16-N和-C端互作及MMD1对其互作的影响
图2 JMJ16-N和-C端互作及MMD1对其互作的影响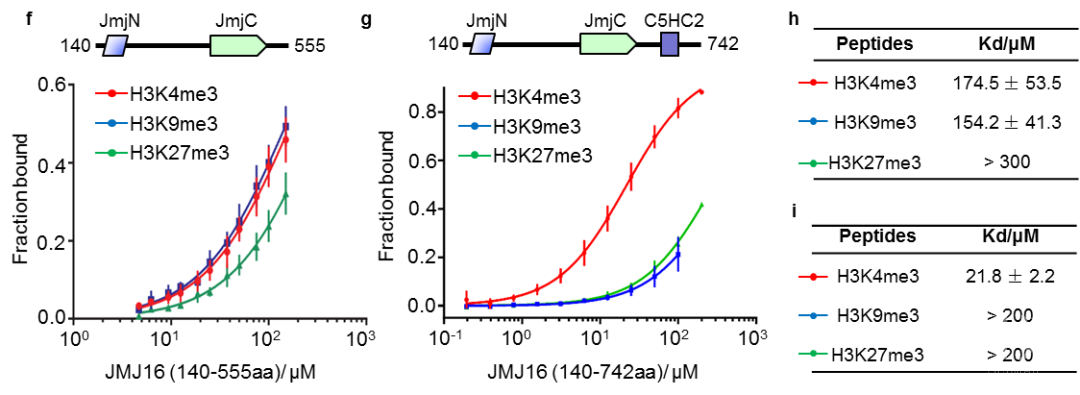 图 3 荧光偏振鉴定 JMJ16酶活结构域及含锌指结构域对H3K4me3和H3K9me3的结合强度
图 3 荧光偏振鉴定 JMJ16酶活结构域及含锌指结构域对H3K4me3和H3K9me3的结合强度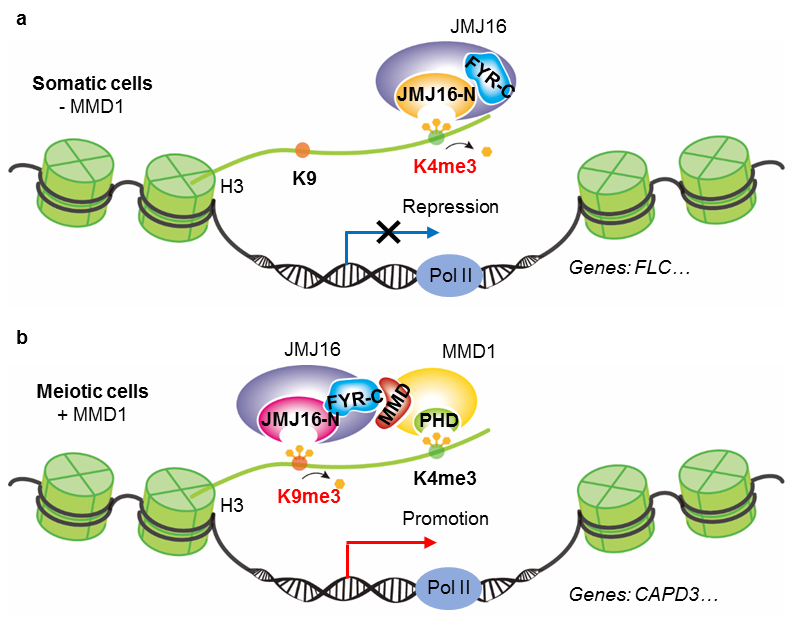 图 4 JMJ16和MMD1在不同细胞微环境中调控基因表达的模型
图 4 JMJ16和MMD1在不同细胞微环境中调控基因表达的模型来源:bioartplants BioArt植物
原文链接:https://mp.weixin.qq.com/s?__biz=MzU3ODY3MDM0NA==&mid=2247496233&idx=1&sn=b1c9680b6471c470089abd7d319e189e&chksm=fd73684eca04e1589bbf05180ed01be3dc57b671d89ce1e70158dca3ed51f1b9acb8f2f664d0#rd
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn

中科院昆明植物研究所通过小垫柳全基因组分析为横断山高山植物多样性起源提供证据
染色体核型分析检查
酵母菌人工染色体载体
ph染色体
无性也能繁衍1800万年! 蠕虫基因测序解密

今日科技话题:地球大气臭氧分布受太阳活动周期影响、 环形染色体重排研究方面取得突破、中国空间站计划实施13次飞行任务
巨染色体
染色体变异与癌症

蛛丝延展性的奥秘

《自然》:端粒T环的保护特性


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号