科技工作者之家
科界APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2019-11-15
来源:土壤观察
导 读
近年来, 极端气候事件发生频率增加, 对陆地生态系统服务功能和人类社会生产生活造成严重影响. 碳循环是驱动陆地生态系统变化的关键过程. 准确理解和评估极端气候事件对陆地生态系统碳循环的影响, 能为人类社会减缓和适应气候变化提供重要科学依据. 文章以干旱、极端降水、极端高温和极端低温为例, 系统总结了极端气候事件对陆地生态系统碳循环的影响及其机理. 已有研究表明, 干旱是当前陆地生态系统碳汇功能的重要胁迫因子, 对生态系统生产力和呼吸都存在压制作用, 但生产力对干旱的敏感性一般高于呼吸对干旱的敏感性, 从而导致陆地生态系统碳汇功能显著削弱, 甚至使之变成碳源. 不同模型对干旱导致的碳循环变化模拟结果差异很大, 显示目前学术界对生态系统碳循环响应干旱机制的认知有限, 尤其是干旱对热带植被生长的影响机制仍存在较大争议. 极端降水事件对生态系统碳循环的影响存在显著区域差异, 一般认为, 极端降水促进干旱地区生态系统碳积累, 却不利于湿润地区生态系统固碳; 但目前对极端降水导致的土壤碳侧向输移和土壤养分流失等间接影响过程的了解十分有限, 致使结果存在很大不确定性. 极端高温和极端低温也通过不同的机制过程影响生态系统碳循环, 尤其值得注意的是其影响程度与这些事件的发生时间存在密切关系, 但这一联系还有待进一步研究. 基于已有认识, 建议未来关于极端气候事件对碳循环影响的研究重点应该是关注其长期效应和不同时间尺度上的作用机理, 并加强基于多数据、多途径的多尺度集成研究.
文/朴世龙1,2,*, 张新平1, 陈安平1, 刘强1, 连旭1, 王旭辉1, 彭书时1, 吴秀臣3(1. 北京大学城市与环境学院, 北京 100871;2. 中国科学院青藏高原研究所, 北京 100101;3. 北京师范大学资源学院, 北京 100875)
来源:中国科学地球科学(2019年第9期)
1 引言
极端气候事件频率、强度和持续时间的增加是全球变化的重要特征之一, 也是人类面临的最严峻挑战之一(Field等, 2012). 近年来, 在全球范围内发生了一系列引起广泛关注的极端气候事件, 例如, 2003年和2010年欧洲地区的高温热浪(Luterbacher等, 2004; Barriopedro等, 2011)、2005年和2010年亚马孙地区的严重干旱(Zeng等, 2008; Lewis等, 2011), 以及2014年初发生在英国的风暴(World Meteorological Organization, 2014, https://www.wmo.int/pages/index_zh.html)等. 尽管地球系统模型预测存在很大的不确定性, 政府间气候变化专门委员会(Intergovernmental Panel on Climate Change, IPCC)第五次评估报告认为, 21世纪全球极端气候事件发生的频率将快速增加且其影响范围呈现扩大趋势(IPCC, 2013). 由于陆地生态系统的适应能力有限, 极端气候事件很可能对陆地生态系统造成更严重、更持久且不可恢复的破坏(Ciais等, 2005). 全面、准确认识和评估极端气候事件对陆地生态系统的影响, 是国内外科学界、政府和公众普遍关注的重大科学问题, 也会为提高人类社会应对和适应气候变化的能力提供重要科学依据.
陆地生态系统碳循环是驱动生态系统变化的关键过程, 并与生态系统水循环、养分循环和生物多样性有着密切的耦合关系. 因此, 在极端气候事件的影响下, 陆地生态系统碳循环的变化将是陆地生态系统响应极端气候事件的综合体现. 另一方面, 陆地生态系统碳循环在减缓大气CO2浓度增加以及全球温度上升方面有着不可替代的作用(Ciais等, 2013). 大量证据表明, 全球陆地生态系统是一个巨大的碳汇. 在过去50年里, 全球陆地生态系统吸收了25~30%的人类活动(主要是化石燃料燃烧和热带雨林砍伐)导致的CO2释放量(Le Quéré等, 2009). 然而, 陆地生态系统碳循环对气候变化十分敏感(Piao等, 2013; Wang等, 2014), 其最明显的特征是, 全球陆地生态系统碳汇具有较大的年际波动, 并决定着大气CO2浓度年增加量的年际波动(图1). 如图1所示, 20世纪80年代以来, 大气CO2浓度年增加量较大的年份, 基本上都发生了较大区域尺度上的极端气候事件, 同时该年份的陆地生态系统固碳能力相对较低. 这意味着, 陆地生态系统碳汇功能对极端气候变化十分敏感. 因此, 陆地生态系统对极端气候的响应研究是全球碳循环研究的重点之一, 也是当前全球变化生态学的核心内容之一.
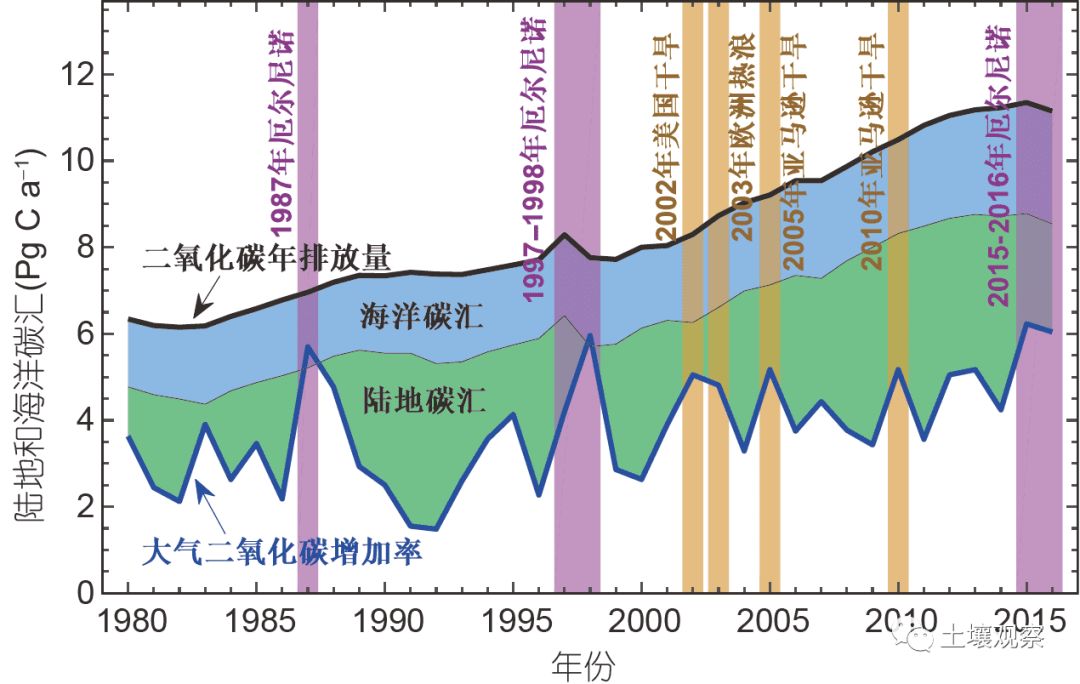
图 11980年以来陆地和海洋碳汇的年际波动
干旱、极端降水、极端高温和极端低温是较为常见的极端气候事件. 因此, 本文拟以上述四类极端气候事件为例, 系统总结极端气候事件对陆地生态系统碳循环影响的表现、范围、过程机理和模型模拟等. 在此基础上, 本文将指出未来这一领域的研究应重点关注的科学问题, 以期抛砖引玉, 激发进一步的研究与讨论. 需要指出的是, 目前研究对不同极端气候事件的深入程度存在很大差异, 因而本文对不同极端气候事件碳循环影响的讨论亦着墨深浅不一.
2 干旱对陆地生态系统碳循环的影响
区域和全球尺度上, 干旱被认为是影响当前陆地生态系统碳汇功能的最主要胁迫因子之一(Ciais等, 2005; Zscheischler等, 2014a). 来自长期定位观测(Knapp和Smith, 2001; Phillips等, 2009)、野外控制实验(Kreyling等, 2008; Jentsch等, 2011; 牟成香等, 2013)、基于涡度相关技术的碳通量观测(Schwalm等, 2010; Yi等, 2012; Huang等, 2013)、大尺度遥感反演(Zhao和Running, 2010; Zscheischler等, 2014a; Xu等, 2011)、模型模拟(Reichstein等, 2007; Zscheischler等, 2014b)等方面的研究均得出比较一致的结论, 即干旱显著削弱生态系统碳汇功能, 甚至使之变成碳源. 例如, 2003年欧洲夏季干旱, 使欧洲生态系统向大气中释放了0.5Pg(petagram, 1015g)的碳, 抵消了此前四年的固碳总量(Ciais等, 2005). Zscheischler等(2014b)利用10种生态系统模型, 估算了过去30年间干旱对全球陆地生态系统碳循环的影响. 其结果表明, 干旱显著降低了全球陆地生态系统碳的净吸收, 导致生态系统碳储量每年净减少0.19Pg, 占全球陆地生态系统碳汇总量的8.3%. 需要指出的是, 不同模型之间的模拟结果差异很大, 而造成这种差异的主要原因在于, 有关碳循环关键过程对干旱响应机制的认识十分有限.
2.1 对植被生产力的影响
植被生产力是生态系统碳汇功能对干旱响应的决定因素之一. 关于干旱对生产力的影响, 学术界已经进行了较为深入的研究. 已有研究表明由于温带大部分地区植被生长受水分显著影响(Nemani等, 2003; Xu等, 2012), 干旱导致温带地区植被生长显著下降; 但关于干旱对热带植被生长的影响, 目前的研究还存在较大争议. Cox等(2000)在Nature杂志上发表文章指出, 未来由于干旱导致的热带森林生产力的下降将使得更多的碳由热带森林向大气释放, 从而进一步加速气候变化. 过去20年间, 这一基于模型模拟的研究结果引起了科学家的广泛关注. 那么, 热带森林生长是否对干旱敏感? 随着未来干旱的加剧, 热带地区生态系统生产力是否大量下降?
自21世纪初以来, 高分辨率遥感数据、长期热带森林样地、生态系统通量观测和更多的区域大气CO2浓度观测数据为科学家更好的研究热带森林碳循环提供了有力支撑, 而2005年和2010年亚马孙地区经历的两次百年一遇的极端干旱(Aragão等, 2007; Lewis等, 2011), 为了解热带森林生长和碳循环对干旱的响应提供了难得的机会. 与Cox等(2000)的模型模拟结果相反, Saleska等(2007)基于MODIS增强植被指数(EVI)数据发现, 在2005年遭遇干旱的亚马孙西南部地区, 植被指数反而显著增加. 部分通量站点的观测数据似乎也表明低降水和低云量导致受辐射强度限制的热带森林在干旱事件发生期间反而吸收了更多CO2(Bonal等, 2008). 如果热带森林被证实能够适应百年一遇的极端干旱并吸收更多的大气CO2, 这将颠覆以往认为干旱导致热带森林向大气释放更多CO2的认识(Cox等, 2004, 2000). 另一方面, 由于云等因素的干扰以及植被指数在热带地区容易“饱和”, 目前遥感数据观测到的热带地区植被指数具有很大的不确定性. 例如, Samanta等(2010)过滤了云和气溶胶等干扰因素之后发现, MODIS增强植被指数(EVI)在2005年极端干旱发生期间并没有较往年增加. 虽然基于遥感植被指数对2005年极端干旱的影响的评估仍然存在较大争议(Asner和Alencar, 2010), 基于长期森林样地的观测则证实了2005年极端干旱导致亚马孙地区森林生产力和碳汇功能显著下降(Phillips等, 2009). 针对2010年亚马孙流域再次发生的极端干旱事件的研究表明, 干旱发生期间, 遥感植被指数(EVI)显著下降(Xu等, 2011). 总之, 从地面到卫星观测, 越来越一致地认为极端干旱事件导致热带生态系统生产力和碳汇功能降低, 虽然其影响强度和机制目前还存在较大争议.
2.2 对生态系统呼吸的影响
生态系统呼吸是仅次于光合作用的陆地生态系统碳通量, 在陆地生态系统碳循环过程研究中处于关键地位. 水分有效性是植物和微生物生长的重要限制因子之一, 干旱条件下, 土壤含水量会显著下降, 而土壤水分的变化会对生态系统呼吸产生重要影响(Liu等, 2009; Vicca等, 2014).
通常情况下, 干旱降低生态系统呼吸, 尽管其响应程度随着不同环境条件而发生变化. 碳通量观测数据表明, 2003年欧洲夏季干旱发生期间, 生态系统生产力下降量大于生态系统碳汇的下降量, 这说明生态系统呼吸也相应减小(Ciais等, 2005). 而针对2010年亚马孙流域严重干旱事件的样地调查也发现, 生态系统呼吸在干旱事件中发生显著下降(Doughty等, 2015).
生态系统呼吸包括植物的自养呼吸和土壤微生物的异养呼吸, 多项研究表明, 干旱条件下, 水分胁迫导致植物生长受限, 从而降低自养呼吸速率; 同时, 由于植物碳底物的供应减少和微生物分解作用下降, 土壤异养呼吸也呈降低趋势(Suseela等, 2012; Zeglin等, 2013; Shi等, 2014; Thakur等, 2018). 另外, 这两种呼吸是完全不同的生物学过程, 对温度等环境因子的敏感性不同, 因此需要分别考虑. 例如, Meir等(2008)分析了亚马孙热带雨林生态系统不同组分(叶、木材、土壤)呼吸对短期干旱事件的响应, 其结果表明, 干旱胁迫下, 生态系统不同组分呼吸均呈下降趋势, 但其下降幅度与机理很不一样. 土壤呼吸对土壤含水量的变化最为敏感, 是受干旱影响最大的组分; 叶片呼吸的降低主要是通过叶面积的下降(即落叶)实现的, 而单位叶面积的呼吸速率大体保持干旱前的水平(Meir等, 2008).
2.3 对群落干扰和生态系统演替的影响
值得注意的是, 干旱不仅显著降低植被生产力, 持续性严重干旱甚至会造成植被大面积死亡(Allen等, 2015), 导致生态系统的衰退或崩溃, 并可能使其最终被别的生态系统类型取代. 例如, 干旱频率的增加导致热带森林树种组成发生改变, 相对更耐旱树种的比例增加(Esquivel-Muelbert等, 2019). 地面调查和遥感数据均表明, 近40年全球范围内广泛出现了树木死亡现象, 而干旱则被认为是导致这一现象的主要原因(Allen等, 2015). 那么, 干旱导致森林死亡的机制是什么? 森林是由于水分亏缺所导致的水力结构损伤失效而“渴”死的(例如, Bréda等, 2006), 还是由于气孔关闭, 光合作用受到抑制导致没有足够的碳维持生命活动而“饿”死的(McDowell等, 2008)? 前者被学界称为“水力传导失效”假说(hydraulic failure), 后者被称为“碳饥饿”假说(carbon starvation). 遗憾的是, 这两种机制对森林死亡的贡献仍不明确, 限制了我们评估过去和预估未来干旱对陆地生态系统碳循环影响. 最近的研究表明, 过去50年热带地区的干旱趋势使得该地区陆地生态系统碳循环发生状态突变(regime shift), 其生态系统碳汇对温度的敏感性增加了近一倍(Wang等, 2014). 然而, 目前国际上应用于生态系统碳循环研究的5种主要过程模型都没有模拟到这一现象.
干旱除了直接减小生态系统生产力、导致树木“水力传导失效”或“碳饥饿”死亡外, 还会通过增加病虫害等干扰间接影响树木死亡和碳源汇功能(图2)(Seidl等, 2018). 例如, 由于干旱导致的松甲虫虫害的增加, 加拿大森林近几十年的死亡率显著增加(Kurz等, 2008). 干旱不仅会增加病虫害爆发的频率、强度和持续时间, 而且可能会改变植物对病虫害的抗性. 但全球和区域尺度上, 干旱通过虫灾间接影响碳循环的研究还很薄弱, 需要进一步加强.
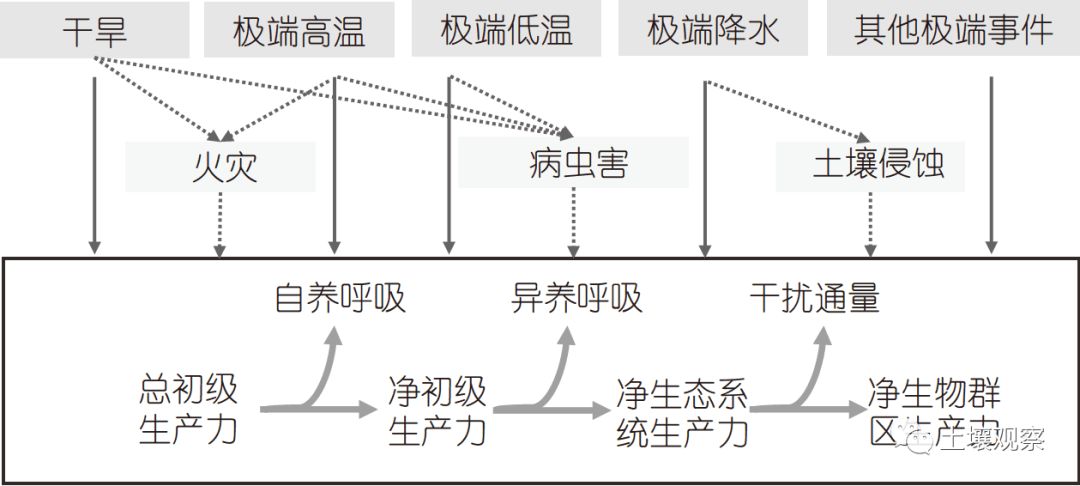
图 2极端气候事件对陆地生态系统碳循环的直接和间接影响
生态系统火灾频率或强度增加也是干旱导致生态系统碳汇功能下降的重要机制之一(图2)(Aragão等, 2008; Brando等, 2014). 已有研究表明, 干旱导致树木含水量降低, 易燃性(flammability)增加, 从而导致森林火灾发生的概率和范围增加(Brando等, 2014). Gatti等(2014)结合大气CO和CO2浓度估算认为, 2010年亚马孙流域火灾所释放到大气中的碳量约为0.51Pg, 这一数值几乎完全解释了同期亚马孙流域的净碳交换量(0.48Pg). 需要指出的是, 虽然卫星数据可以较好地估计过火面积, 然而由于可燃物(冠层树木、林下植被和凋落物)储量的不确定性, 生态系统火灾导致的CO2排放量仍然存在较大不确定性. 利用卫星获取的燃烧面积、大气CO2和CO浓度观测以及地面清查资料相结合的手段, 将有助于我们进一步定量估计干旱导致的火灾对生态系统碳循环的影响.
3 极端降水对陆地生态系统碳循环的影响
在气候变暖的背景下, 极端降水发生的频率和强度都很可能增加(IPCC, 2013), 但有关极端降水对生态系统碳循环的影响研究还比较薄弱. 通常, 极端降水对生态系统碳循环的影响主要取决于极端降水发生时生态系统的水分条件(即是否受水分限制)(Knapp等, 2008; Zeppel等, 2014)以及极端降水的发生时间(Fay等, 2011; Zeppel等, 2014). 已有研究表明, 在干旱地区, 极端降水导致土壤水分增加, 从而提高生态系统生产力和碳积累; 而在湿润地区, 极端降水不利于生态系统生产力和固碳功能(Knapp等, 2002, 2008; Heisler-White等, 2009; Fay等, 2011; Zeppel等, 2014), 尤其在长时间淹水状态下, 植被根系甚至严重缺氧、呼吸减少, 并可能最终引起植被死亡(Kramer等, 2008). 另外, 不同季节的极端降水事件对生态系统碳循环的影响也有所差别. 例如, 春季和夏季的极端降水对干旱地区草地生态系统生产力的促进作用远大于秋季和冬季极端降水的作用(Zeppel等, 2014). 值得注意的是, 目前有关生态系统碳循环对极端降水的响应研究大部分是通过生态系统的增水控制实验途径进行的, 且集中在草地生态系统, 因此迫切需要加强对森林生态系统及其他植被类型的研究.
另一方面, 极端降水常伴随着龙卷风或风暴, 以单次降雨强度增大为特征, 因此极端降水事件的增多可能会加剧土壤侵蚀, 由此加剧土壤有机碳的剥蚀、迁移和沉积过程, 因而也会改变陆地碳源汇的大小和空间分布(例如, Van Oost等, 2007; Frank等, 2015). 近些年来, 尽管土壤侵蚀导致碳的侧向通量在全球碳收支平衡中的重要作用已被逐渐认知(例如, Van Oost等, 2007, 2012; Hoffmann等, 2013), 但这一过程的净作用究竟表现为碳汇(Stallard, 1998; Van Oost等, 2007, 2012), 还是碳源(Lal, 2003) 目前还存在较大分歧. 一般认为, 土壤侵蚀所引发的土壤碳侧向运移是增强还是减弱陆地碳源汇大小取决于土壤碳迁移的路径和最后沉积的位置(Yue等, 2016): 如果被沉积土壤碳的周转周期大于原来生态系统的土壤碳周转周期, 则有利于碳的积累, 反之则会加剧碳的释放(Van Oost等, 2007; Yue等, 2016). 另外, 土壤侵蚀会导致土壤养分的流失, 从而间接影响生态系统生产力和碳源汇功能. 最近, Zscheischler等(2014b)利用多个模型的模拟结果阐明, 全球尺度上极端降水引起的生态系统生产力的增加量大于同时期生态系统呼吸的增加量, 从而提高了陆地生态系统的固碳能力, 但其所用的模型并未考虑土壤碳的侧向输移和土壤养分流失对生态系统碳循环的间接影响, 因而其结果还具有很大的不确定性.
4 极端高温对生态系统碳循环的影响
20世纪中期以来, 全球许多地区高温热浪事件频发, 且持续时间和强度增加(Della-Marta等, 2007; Perkins等, 2012; IPCC, 2013), 热浪对生态系统碳循环的影响及其未来变化趋势受到广泛关注. 在极端高温胁迫下, 植被光合作用减弱甚至停滞, 呼吸增加(Salvucci和Crafts-Brandner, 2004), 且呼吸的增加量大于植被生产力的减少量, 从而加剧生态系统碳汇功能下降. 这一机制与干旱导致生态系统碳汇减少的原因恰好相反(von Buttlar等, 2018). 例如, 在2010年俄罗斯西部的高温热浪事件中, 生态系统生产力下降, 同时植被呼吸增加(Bastos等, 2014). 在自然条件下, 极端高温的发生往往伴随着降水的减少和空气相对湿度的降低(De Boeck等, 2010), 这给我们准确认识极端高温对碳循环的影响带来了一定的不确定性. 相对于单一的热浪事件, 极端高温与干旱的协同作用会加剧植被生产力下降, 从而加剧生态系统碳的流失(von Buttlar等, 2018). 而极端高温与干旱对呼吸作用的协同影响相对较小, 这主要是由于热浪引起的植被呼吸作用增加与干旱引起的植被呼吸作用减少相互抵消(von Buttlar等, 2018).
极端高温对生态系统碳循环影响的大小不仅取决于热浪强度, 还与热浪的发生时间有密切关系(Sippel等, 2016; Perkins-Kirkpatrick和Gibson, 2017; Sippel等, 2018). 过去大部分关于高温热浪事件对碳循环影响的研究都是围绕夏季高温热浪事件来开展的, 而已有研究表明, 不同季节发生的高温热浪事件对碳循环的影响是不同的. 例如, 不同于夏季高温热浪通常降低生态系统固碳, 春季的高温热浪可能通过提前春季物候促进生态系统碳的净吸收(De Boeck等, 2011). 这一结论也得到欧洲长期碳通量观测数据的支持. Delpierre等(2009)利用欧洲碳通量观测网数据分析了2007年异常暖春对碳循环的影响, 发现在不同的生态系统中暖春事件都导致生产力和呼吸增加, 而且生产力的增加量高于呼吸的增加量, 从而有利于春季碳吸收. 然而, 最近的研究表明, 尽管春季高温有利于生态系统春季碳吸收, 但由于春季植被生长增加导致的植被蒸腾的提高, 间接引起夏季植被的水分亏缺, 从而降低夏季生态系统碳汇能力, 使生态系统年碳吸收量变化不大(Wolf等, 2016; Buermann等, 2013).
5 极端低温对生态系统碳循环的影响
相对于极端高温, 人们对于极端低温事件的关注较少, 这与全球变化背景下极端低温出现的概率低于高温热浪事件有关(Field等, 2012). 研究结果表明, 虽然气候变暖减少霜冻事件(即frost days, 日最低温度低于0℃的天数)的发生(IPCC, 2013), 但植被生长期间发生霜冻事件的天数不一定减少, 甚至有些地方(比如: 欧洲)表现为显著增加(Liu等, 2018). 这主要与气候变暖导致的植被生长季长度延长有关. 来自地面观测和遥感数据一致表明, 近30年来, 气候变暖导致北半球生长季长度快速变化, 表现为春季物候提前和秋季物候延迟(Fu等, 2015; Liu等, 2016).
已有研究表明, 植被生长季延长是气候变暖促进北半球植被生长的重要机制(White等, 1999; Piao等, 2007), 然而生长季初期和末期极端低温事件的增加可能在一定程度上抵消生长季延长对生产力增加的贡献. 植被生长期间发生的极端低温会导致植被发育缓慢(Polle等, 1996; Dittmar等, 2006), 而冰冻引起的细胞脱水则会造成植物组织(比如, 花蕾、叶片等)冻害, 甚至死亡(Inouye, 2000, 2008; Snyder和Melo-Abreu, 2005). 例如, Dittmar等(2006)基于德国南部的树轮数据分析指出, 极端低温事件导致树轮的径向生长降低了90%. Vanoni等(2016)通过分析瑞士14个自然保护区内9个温带树种的树轮数据发现, 春季晚期的极端低温事件导致绝大部分树种生长快速减慢. 区域尺度上, Hufkens等(2012)指出2010年美国东北部春寒导致该地区高海拔森林的年总初级生产力下降了7~14%. 这些研究结果暗示着极端低温对生态系统碳循环的影响不可小觑, 需要加强相关研究.
6 存在问题与未来研究方向
综上所述, 近几十年来, 极端气候事件的频繁发生对全球和区域碳循环有着不容忽视的影响. 科学界利用不同来源的观测数据以及模型模拟手段, 对极端气候事件影响生态系统碳循环的表现、范围以及过程机理等进行了大量研究, 得到了一些重要结论. 然而, 目前我们对这一复杂问题的认识还存在诸多不足, 主要表现在如下几个方面: (1) 对极端气候影响碳循环的机制认识还存在较大的不确定性; (2) 对极端气候影响下生态系统碳循环的长期动态和生态系统功能恢复过程了解有限; (3) 在极端气候影响下的生态系统碳循环研究中, 需要进一步加强多源数据与模型模拟的结合. 这些是未来研究应该着重关注的问题.
6.1 极端气候影响碳循环的机制研究
宏观尺度上, 植被在长期的适应演化中与环境动态达到了一定的平衡. 而极端气候无论是干旱、极端降水、高温热浪, 还是极端低温, 都偏离植被所长期适应的环境, 从而抑制植被生长甚至导致植被死亡, 并进一步影响区域乃至全球陆地生态系统碳源汇的大小和方向. 另一方面, 植被对极端气候也有一定程度的适应和抵抗能力. 当极端气候的强度和频度没有超出植物的忍耐限度时, 极端气候事件通常显著降低植物固定大气CO2的速率和能力, 即植被生产力, 并可能加快生态系统代谢, 从而降低生态系统碳汇功能甚至使生态系统由碳汇转变成碳源. 当极端气候的强度和频率超出植物的忍耐限度时, 极端气候事件会导致植被的大面积死亡. 极端气候导致的植被生产力快速下降, 以及死亡植被的长期分解过程, 均是生态系统碳排放的重要来源. 总之, 极端气候对生态系统碳平衡通常是一个负面的影响. 准确定量描述这一影响, 有赖于科学家们对极端气候影响生态系统碳循环机制的深入理解, 包括如何提高对植被生长、死亡, 以及抵抗极端气候事件的机制的准确描述. 然而, 过去的工作大多侧重于现象描述, 结合理论模型对其背后的驱动机制的深入分析尚较为缺乏.
植被死亡是极端气候事件影响生态系统碳循环的重要途经. 如前文所述, 以干旱为例, 其导致植物死亡的机制存在“水力传导失效”和“碳饥饿”两种主要假说(Bréda等, 2006; McDowell等, 2008; Anderegg等, 2013; Adams等, 2017; Choat等, 2018; Hartmann等, 2018), 然而, 针对这两种主要假说的研究还存在很多争议和不清楚的地方. 例如, 通常认为, 干旱胁迫下木质部发生栓塞(embolism)以及导管水势(xylem water potential)下降是导致树木水分亏缺乃至死亡的主要因素(Hoffmann等, 2011; Urli等, 2013; Anderegg等, 2016; Choat等, 2018; Hartmann等, 2018), 但导致树木死亡的木质部栓塞或导管水势的阈值目前还较难定量测算(Choat等, 2018; Hartmann等, 2018). 另外, 这个阈值在不同区域、不同物种之间可能存在显著差异, 这些差异增加了模型模拟的复杂度. 技术进步如遥感、X-射线扫描、离心分析等为监测和检测树木气穴和栓塞以及由此导致的树木死亡等提供了新的可能途径(Brodribb等, 2016; Choat等, 2016; Rodriguez-Dominguez等, 2018), 但这些技术目前还比较昂贵, 其广泛适用性也有待进一步研究. 许多关于干旱导致树木死亡的实验(Brodribb和Cochard, 2009; Urli等, 2013)是利用幼龄树完成的, 尚缺乏成熟树木的长期干旱模拟实验. 干旱导致的树木水力传导下降和碳饥饿机制也可能存在交互关联, 但这些关系我们目前尚不清楚. 其他问题包括: 干旱与其他环境过程如何协同影响植物生理生态过程, 从而影响植物个体死亡和生态系统碳循环? 如何在陆面过程模型中准确表达这一机制? 同时, 如何将个体水平的机理描述转换到区域乃至全球尺度上, 尤其针对多树种、多气候区的影响分析? 除了干旱以外, 其他极端气候事件如飓风、极端低温和降雪等也可能导致一定程度的植被死亡, 但目前的研究对极端气候导致的植被死亡多是以概率模型进行描述, 缺乏进一步的机理研究和过程模拟.
生态系统对极端气候事件存在一定程度的抵抗力, 准确理解和描述这一抵抗机制及其限制也是提高模型模拟生态系统碳循环响应极端气候能力的一个重要因素. 一般而言, 植被生长对干旱的抵抗力与植物功能属性间存在密切关系, 植被结构复杂性(如物种多样性)对生态系统应对极端事件的抵抗力亦有显著影响(Isbell等, 2015), 生态系统对不同类型极端气候事件的抵抗力也存在显著分异. 例如, 美国中部草原生态系统对极端高温表现出较高的抵抗力, 但对极端干旱的抵抗力较低(Hoover等, 2014). 然而, 我们对上述现象的理解仍面临挑战: (1) 针对不同气候区、不同植被功能类群对极端气候事件抵抗力的系统研究仍十分缺乏; (2) 生态系统对不同类型极端气候(如连续型干旱、离散型干旱、极端降雪等)及不同极端气候事件特征(如干旱强度、持续时间等)抵抗力的对比研究不足; (3) 缺乏生态系统对极端气候事件抵抗力内在机理规律(如生理生态过程、植物功能属性调控等)的系统研究.
6.2 极端气候对生态系统碳循环的长期影响与生态系统的恢复力研究
极端气候事件对生态系统的影响可能存在一定的“遗产”效应(legacy effect), 即在极端气候事件结束以后, 由于植物生长的部分环境限制因素(如土壤水分含量)及植被生理生态功能不能及时恢复, 生态系统碳循环功能仍将继续受损. 例如, 干旱对树木生长的影响可能会持续1~4年, 这一遗产效应在温带和寒带森林生态系统中广泛存在(Anderegg等, 2015). 干旱遗产效应的持续时间和大小与植被的水分传导属性(Anderegg等, 2015)及干旱发生的季节(Huang等, 2018)都可能有一定的关系. 深入理解和准确模拟这一遗产效应机制将有助于提高我们对极端气候事件对生态系统碳循环影响的认知.
除了遗产效应, 评估极端气候对陆地碳循环的长期影响亦应包括生态系统的自我恢复过程. 然而, 目前研究焦点大都放在了碳循环过程对极端气候响应的敏感性方面, 而较少关注极端气候事件发生后的生态系统碳汇功能的恢复力问题(Haddad等, 2002; Van Ruijven和Berendse, 2010; Hoover等, 2014). 研究恢复力之所以重要, 是因为生态系统的快速恢复能部分抵消极端气候事件对生态系统碳汇功能的负面影响(例如, Amiro等, 2010). 恢复力的强弱不仅与生态系统的属性有关, 而且还取决于极端气候事件的性质和强度. 例如, 2010年发生在中国西南边陲的春旱, 虽然降低了大部分西南地区春季和初夏的植被生产力, 但是这种负面效应并没有延续到夏末和秋季的植被生长(Zhang等, 2012; Li等, 2019). 通过研究恢复力, 并结合碳循环过程对极端事件响应的敏感性分析, 能让我们综合评估碳循环对极端气候事件响应的韧性和脆弱性, 为科学应对极端气候事件、实现趋利避害提供依据.
未来关于生态系统恢复力的研究应在以下几个方面予以特别关注: (1) 极端气候事件与其他干扰(如病虫害)相互作用对生态系统恢复力的综合研究(He等, 2017). 最近的研究表明, 考虑对外来虫灾的管理, 能显著缩短森林碳汇的恢复时间(Seidl等, 2018). (2) 生态系统恢复力在短时间尺度及长时间尺度上的分异特征如何? 最新的研究表明, 森林更新导致的森林结构的长时期变化对森林生态系统的恢复力具有重要影响. (3) 生态系统对极端气候事件或干扰的恢复力的变化动态如何? (4) 生态系统恢复力与生态系统响应敏感性的多样性(sensitivity diversity)之间存在什么样的内在关联?
6.3 多源数据融合与极端气候事件影响下生态系统碳循环模型模拟研究
地面样方、涡度相关通量塔以及大尺度遥感观测可以为我们研究观测时期内极端气候事件如何影响生态系统生产力和碳源汇功能提供大量观测数据. 但是极端气候事件发生的概率一般很小, 而已有的野外控制实验和基于涡度相关技术的碳通量观测的时间跨度相对较短, 这在一定程度上限制了我们深入理解生态系统碳循环对极端事件的响应机制及其恢复能力. 树木年轮因较为有效地反映了历史时期森林植被的逐年生长状况, 常常被用来指示植被生产力在长时间尺度上的年际波动, 这为我们在更长的时间尺度上研究极端气候对生态系统生产力的影响提供了宝贵资料(Anderegg等, 2015; Huang等, 2018). 因此, 基于不同数据源探讨极端事件对生态系统碳循环影响的不同过程, 有助于我们更全面深入地了解整个碳循环对极端气候事件的响应.
另一方面, 利用过程模型模拟生态系统碳循环是理解和预测生态系统碳循环如何响应极端气候事件的重要方法. 然而, 如何整合长期实验观测网络及多尺度遥感数据, 改进陆面模式中树木死亡机理过程的刻画, 进而系统地理解生态系统碳循环过程对极端气候事件的响应, 是目前所面临的一个重大挑战. 目前, 极端气候影响下的生态系统碳循环模拟研究结果中存在很大的不确定性. 例如, Xiao等(2009)利用生态系统模型(TEM)研究了20世纪不同时期的极端干旱事件对中国陆地生态系统碳源汇功能的影响, 发现绝大部分干旱事件都导致生产力下降量大于生态系统呼吸的下降量, 以至于陆地生态系统从碳汇变成碳源. 然而, 最近的一项基于BEPS模型的研究(Liu等, 2014)却表明, 干旱导致中国陆地生态系统成为碳源的机制在不同的年份有所差别: 在2001年和2011年是由于干旱造成的生产力下降量高于呼吸下降量; 而2009年则是干旱促进生态系统呼吸量和降低生产力. 由此可见, 目前模型研究的结果高度依赖于具体模型的构造和对关键生态过程的描述, 因此迫切需要发展准确详细刻画生态系统过程的机理模型, 结合多源观测数据、控制实验和模型-数据融合方法, 优化模型参数, 并采取多模型的比较模拟研究. 针对极端气候事件对生态系统的影响, 图3提供了一个基于多源数据融合和陆面过程模型模拟的综合研究框架. 遥感、树木年轮、地面观测、控制实验等多源数据为气候、植被、CO2数据等驱动下的陆面生态系统模型提供了参数优化和结果检验; 同时, 通过对多源数据和模型模拟结果的分析, 有利于揭示不同尺度陆地生态系统碳平衡对极端气候事件的响应机制. 发展这种基于多数据、多方法的多尺度集成框架, 将能够更准确地模拟和预测不同极端气候事件下的生态系统碳循环过程, 从而更好地理解并评估生态系统功能对极端气候事件的敏感性、脆弱性以及可能的临界阈值, 为应对极端气候事件的科学决策提供依据.
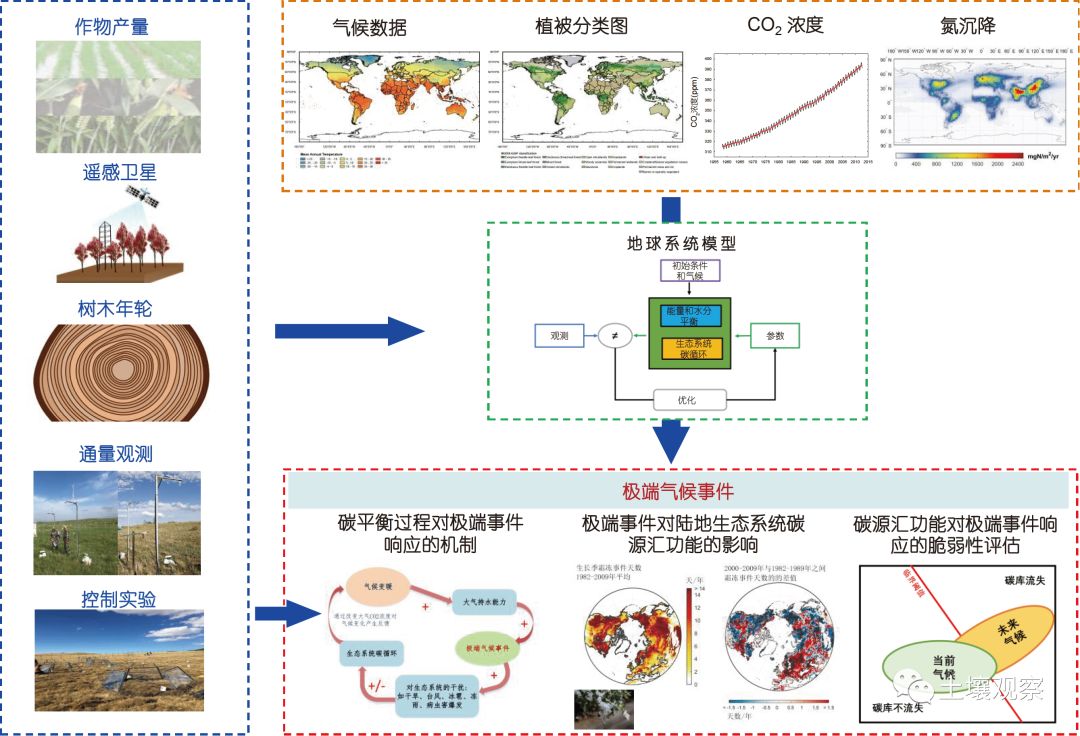
图 3多源数据和多种方法综合研究极端气候事件对陆地生态系统影响的框架图
7 结论
近年来不同极端气候事件的频发给生态系统和人类社会带来了很大影响. 本文基于已有研究, 系统分析了干旱、极端降水、极端高温和极端低温四类极端气候事件对陆地生态系统碳循环的影响及其作用机制. 目前研究普遍表明, 这四类极端气候事件通常会胁迫植被生长, 从而削弱陆地生态系统碳汇功能, 甚至使之变成碳源. 但是, 不同极端气候事件对生态系统碳循环功能的影响以及不同生态系统的响应机理存在显著差异, 不同研究的结论尚存在较大争议和不确定性. 这种不确定性给模型模拟和预测极端气候影响下的陆地生态系统碳循环带来了较大困难. 因此, 作者建议未来研究中加强极端气候事件对碳循环影响的观测和机理研究, 尤其需要重点关注极端气候事件的长期生态系统效应和在不同时间尺度上的作用机理, 并发展基于多数据、多途径的多尺度集成研究, 以提高模型模拟和预测未来气候变化下的生态系统碳循环响应的能力.
致谢
感谢沈泽昊教授、汪涛研究员、刘永稳博士、李相怡、王锴在写作过程中的帮助.
来源:turangguancha 土壤观察
原文链接:http://mp.weixin.qq.com/s?__biz=MzA3MDMwNTExNg==&mid=2659319053&idx=1&sn=f3968343d7e502d703bcde4b4e19f7d3&chksm=844b893eb33c002831ff830c9634fabe38c9757b1b916305dae8c7a374363e3c8529a6ad7dd6&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn
生态系统碳通量测定
生态系统发育
气候变化:树木年轮为青藏高原温差缩小带来启示
草原生态系统
生态系统碳循环

中国环境科学学会王志华秘书长出席第九届国际生态系统服务大会开幕式并致辞

气候变化环境下热带树木不好监测?新研究来助力!
生态系统多样性

生态系统对人类的福祉
生态系统生产力


科界APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号