科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2019-12-04
来源:BioArt
责编 | 兮
基因表达是一个对所有有机体都非常重要的生物过程。有机体可以通过对RNA的加工实现对基因表达的调控。真核生物中,几乎所有的mRNA前体(pre-mRNAs)在从细胞核运输到细胞质进行翻译之前,必须经过额外的加工才能成为成熟的mRNA【1】。这些加工过程包括5’端加帽子,3’端加多腺苷酸的尾巴(poly(A) tail)以及剪接(splicing)。其中3’端加尾是基因调控的主要手段之一【2】,该过程包括两个步骤:1)对pre-mRNA 3’端的特异性位点进行切割;2)在该位点添加poly(A)尾巴。有趣的是,这两个步骤的完成需要体内超过20个蛋白质的参与。这些蛋白质在真核生物中高度保守,它们形成不同的功能模块,彼此相互作用形成庞大的加工机器【2】。
基于对pre-mRNA 3’端加工的三个主要的序列元件的识别,该过程涉及三个重要的蛋白模块:哺乳动物聚腺苷酸化信号因子mPSF (mammalian polyadenylation specificity factor,识别聚腺苷酸化信号AAUAAA),哺乳动物切割因子mCF (mammalian cleavage factor,识别切割位点CA)和切割促进因子CstF (cleavage stimulation factor,识别GU-rich序列)。虽然对3’端加工的生化研究已有40多年的历史,但科学家对这一基本生命过程的具体分子机制依然知之甚少。
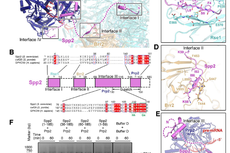
2017年末,美国哥伦比亚大学童亮研究组和洛克菲勒大学Thomas Walz研究组合作解析了蛋白模块mPSF识别聚腺苷酸化信号AAUAAA的冷冻电镜(Cryo-EM)结构,阐明RNA的识别需要WDR33和CPSF30的共同参与, 脚手架蛋白CPSF160起到对WDR33和CPSF30预组装的作用【3】。
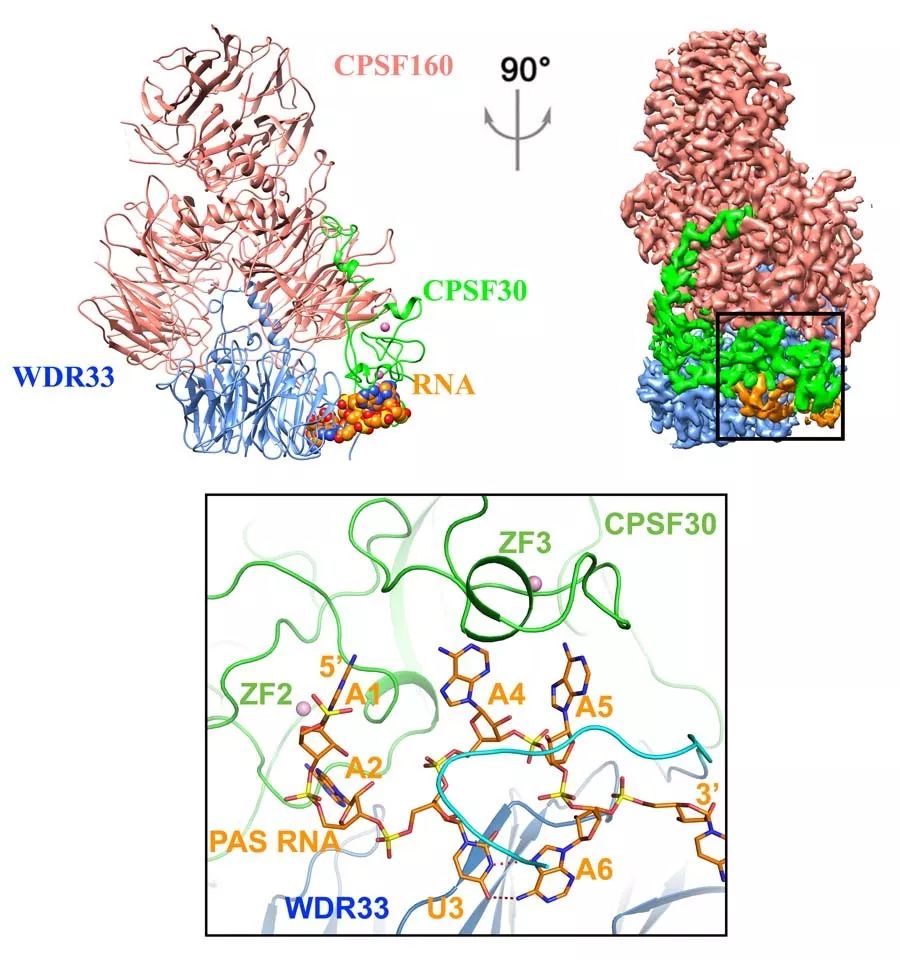
图1 mPSF识别聚腺苷酸化信号AAUAAA的分子机制
为了进一步揭开pre-mRNA的3’端加工机制,童亮研究组和Thomas Walz研究组于2019年12月3日在Molecular Cell在线发表论文,题为:Structural insights into the human pre-mRNA 3'-end processing machinery。

作者分别对蛋白模块mCF, CstF, mPSF-mCF, mPSF-CstF以及mPSF-mCF-CstF进行了深入的结构和功能研究。首先研究人员发现mCF自身的结构处于高度的不均一状态。mCF由核酸内切酶CPSF73,其同源蛋白CPSF100(功能未知)和脚手架蛋白Symplekin组成【4】。这三个蛋白之间的相对位置一直处于灵活多变的状态。即使mPSF识别AAUAAA后招募mCF的结合,形成mPSF-mCF复合物,依然不能稳定mCF的结构。除了mCF自身的不均一性以外,研究发现mCF整体的位置相对于mPSF 也是高度动态的,双重不稳定性阻碍了对mPSF-mCF复合物的高分辨结构解析。
经过研究人员的努力,收集大量冷冻电镜数据并采取对复合物中mPSF和mCF分别处理的策略,分别解析了mPSF (3 Å) 和mCF (7.4 Å) 的电镜结构。有趣的是,研究人员发现与之前mPSF-AAUAAA结构相比,在mPSF的结构中出现了一段新的多肽链。该新的密度含27个氨基酸,横跨40 Å的距离。这27个氨基酸可能来自于处于高度动态的mCF 模块。经过多次尝试,并结合生化实验验证,研究人员发现该多肽位于CPSF100中一段长达108个氨基酸的高度亲水的loop区域。研究人员将这段肽命名为 PIM (PSF interaction motif), PIM与CPSF160和 WDR33分别形成疏水相互作用。PIM 突变体或缺失PIM将不能再形成mPSF-mCF复合物,说明mCF通过处于loop区域的PIM拴在mPSF附近,这刚好解释了mCF整体相对于mPSF 高度动态的原因。
研究人员进一步加入蛋白模块CstF (即CstF77-CstF64-CstF50),发现CstF64和CstF50蛋白相对于CstF77的位置高度动态。而mPSF-mCF-CstF复合物(约800KD)由于其多重不稳定性而无法进行高分辨结构的解析。研究人员经过努力解析了mPSF-CstF77复合物3.6 Å的电镜结构,并指出CstF77的结合同时需要CPSF160和 WDR33。
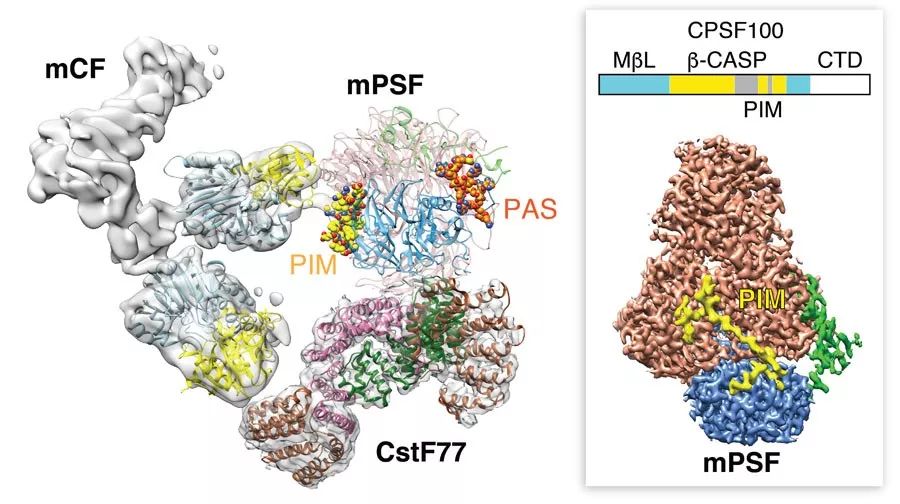
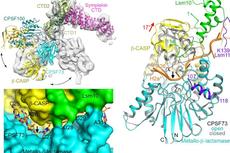
图2. 人类pre-mRNA 3’端加工机器的三维组装模型
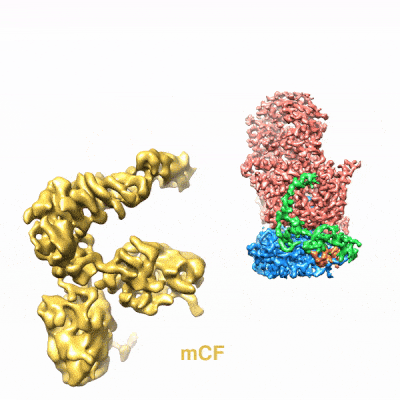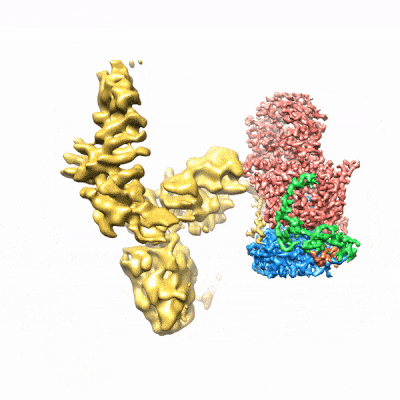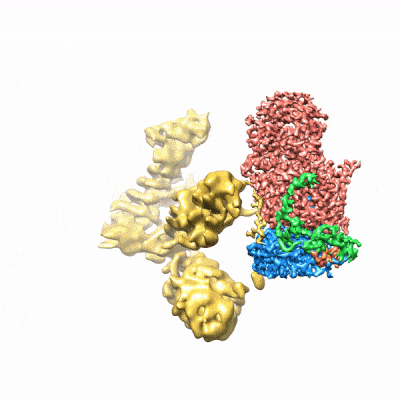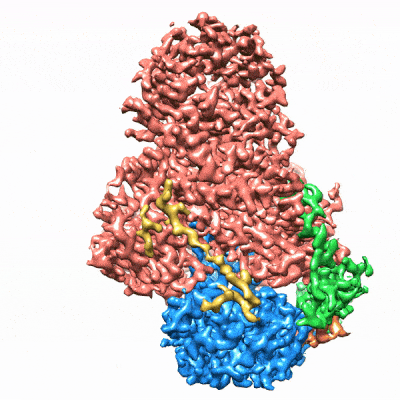
图3. mCF(类三叶草结构)在mPSF周围运动)
综上,该论文通过对人类pre-mRNA 3’端加工机器mPSF-mCF和 mPSF-CstF模块的结构和功能研究,阐明mCF和CstF的不均一性,首次提供了该机器的三维组装模式。并结合之前对mPSF的研究,发现CPSF160-WDR33复合物是该加工机器的核心/中心,招募CPSF30, RNA、mCF和 CstF结合于该核心的不同区域。
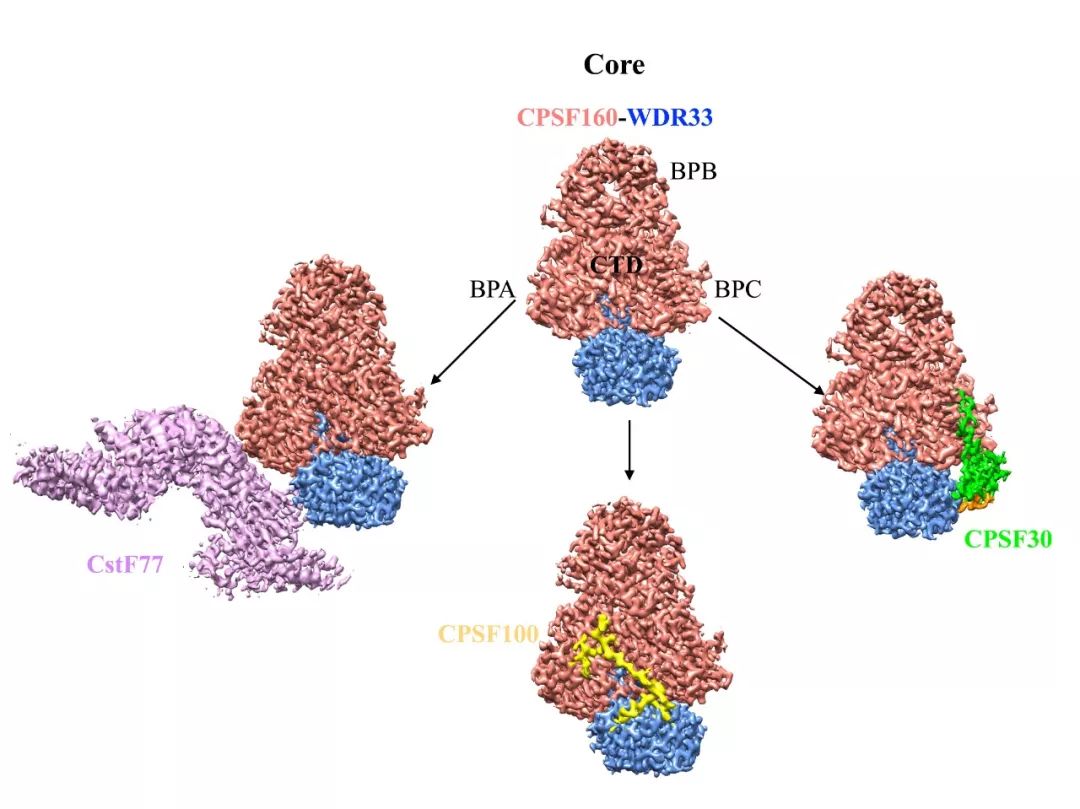
图4. CPSF160-WDR33复合物作为该加工机器的核心,招募相关的蛋白模块
更有趣的是,研究人员发现在该结构中核酸内切酶CPSF73的活性位点距AAUAAA信号序列太远,大于120Å,而已知AAUAAA与切割位点CA之间一般存在10~30个核苷酸的距离【5】。所以mCF 必须进行一系列的结构重排,以实现对RNA的切割。研究人员认为mCF在该加工机器中的双重不稳定性很可能正是其对不同RNA加工得以实现的原因。mCF的激活机制将是下一个值得期待解决的谜题。
美国哥伦比亚大学童亮研究组的孙亚东博士和洛克菲勒大学 Thomas Walz研究组的张一小博士为该论文的共同第一作者。
参考文献
1, Xiang, K., Tong, L., and Manley, J.L. (2014). Delineating the structural blueprint of the pre-mRNA 3' end processing machinery. Mol Cell Biol 34, 1894-1910.
2, Tian B, Manley JL. Alternative cleavage and polyadenylation: the long and short of it. Trends Biochem Sci 2013; 38:312-20.
3, Sun, Y., Zhang, Y., Hamilton, K., Manley, J.L., Shi, Y., Walz, T., and Tong, L. (2018). Molecular basis for the recognition of the human AAUAAA polyadenylation signal. Proc Natl Acad Sci USA 115, E1419-E1428. (Epub Dec. 5, (2017))
4, Mandel, C.R., Kaneko, S., Zhang, H., Gebauer, D., Vethantham, V., Manley, J.L., and Tong, L. (2006). Polyadenylation factor CPSF-73 is the pre-mRNA 3'-end-processing endonuclease. Nature 444, 953-956.
5, Tian, B., Hu, J., Zhang, H., and Lutz, C.S. (2005). A large-scale analysis of mRNA polyadenylation of human and mouse genes. Nucl Acid Res 33, 201-212.
来源:BioGossip BioArt
原文链接:http://mp.weixin.qq.com/s?__biz=MzA3MzQyNjY1MQ==&mid=2652478488&idx=3&sn=c9e4a9d6c1b75d9f3fce0db706be7cb1&chksm=84e20facb39586ba603598ad44112da49f4e654af6e703b2e4edc00e89541b646bd4c91c9df2&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号