科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2019-12-16
来源:宏基因组
细菌和真菌是共存于自然界的两类主要植物致病菌。总体来说,植物周边的细菌数量远多于真菌【1】,而真菌对植物的危害则要甚于细菌【2】。古老的壶菌门真菌已经能够侵染海洋中陆地植物的祖先——藻类【3】,而如今全球作物在生产或贮存中的损失大约有30%由真菌造成【4】。高等植物如拟南芥可通过细胞表面的免疫受体识别细菌或真菌的保守分子(比如细菌的鞭毛多肽flg22和延伸因子多肽elf18以及真菌的细胞壁多糖几丁质)而激活相应的抗性,但较低等的苔藓仅能够识别几丁质【5】。基于进化过程中真菌对植物的巨大威胁,植物是否能够通过识别数量较多的细菌来对数量较少的真菌进行预警呢?
12月11日,中山大学生命科学学院李剑峰教授团队在Cell Host & Microbe发表了题为Cross-microbial protection via priming a conserved immune co-receptor through juxtamembrane phosphorylation in plants 的研究论文。该研究揭示了拟南芥对病原细菌的识别会诱导其几丁质共受体CERK1发生胞内近膜区磷酸化,从而使该受体进入持续48小时以上的“警戒”状态,期间能够更快更持久地转导几丁质信号并获得增强的真菌抗性。这一现象使人联想起淝水之战中前秦的苻坚被东晋军队惨败后“草木皆兵”的典故。值得一提的是,Cell Host & Microbe 同期刊发了日本近畿大学(Kindai University)Tsutomu Kawasaki教授撰写的题为PRR cross-talk jump starts plant immunity 的点评文章。


该研究首先发现真菌(灰霉菌)与细菌(丁香假单胞菌或青枯劳尔氏菌)共侵染拟南芥会使植株出现更强的真菌抗性,且该抗性依赖于拟南芥的几丁质共受体CERK1。令人意外的是,上述细菌侵染或细菌flg22或elf18刺激,均会诱导CERK1胞内近膜区 (juxtamembrane) 的三个氨基酸位点发生磷酸化。后续生化和遗传证据表明,flg22和elf18的共受体BAK1介导了对CERK1的磷酸化。有趣的是,BAK1虽然也是植物激素油菜素内酯的共受体,但该激素激活的BAK1不会磷酸化CERK1。进一步研究发现,近膜区三个位点模拟磷酸化缺失突变的CERK1会使cerk1突变体回补植株失去细菌诱导的真菌抗性,而三个位点模拟持续磷酸化突变即使无细菌诱导也能使植株产生对几丁质更快更持久的免疫应答和对多种真菌的更强抗性。重要的是,在无真菌侵染时,这种真菌抗性增强的植株生长正常,而转录组分析也未见免疫基因异常激活,提示细菌诱导的CERK1近膜区磷酸化仅使该受体进入“警戒”状态而非激活状态。的确,与flg22和elf18不同,真菌几丁质诱导CERK1整个胞内区(包括近膜区和激酶区)发生彻底磷酸化并激活。上述CERK1介导的由细菌诱导的真菌抗性只存在于十字花科拟南芥及近源三个属的植物中,提示这些植物的共同祖先首次进化出这种免疫预警机制。
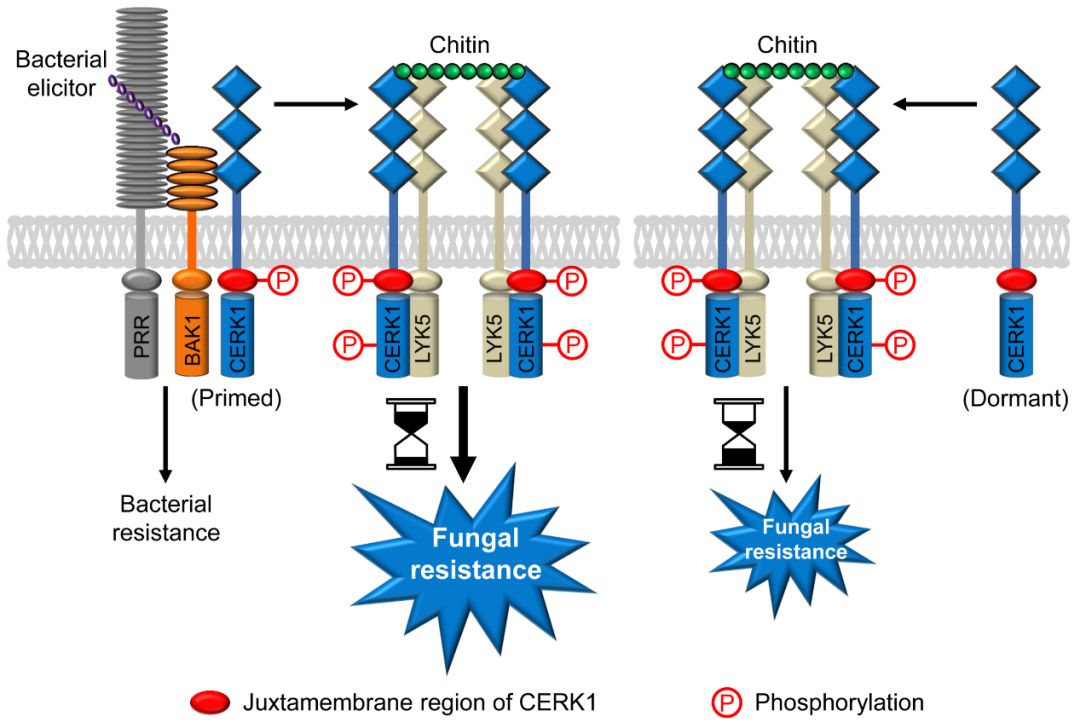
综上,该研究首次发现拟南芥BAK1和CERK1分别作为共受体的两条过去认为彼此独立的免疫信号转导通路存在信息流动(crosstalk),并因此产生细菌诱导的真菌抗性。同时,该研究首次发现植物受体激酶可通过近膜区磷酸化进入一种介于“失活”和“激活”的中间状态,即“警戒”(primed)状态。值得注意的是,茄科植物本氏烟的CERK1因为近膜区没有对应拟南芥CERK1的可磷酸化位点,不能响应flg22进入警戒状态,而拟南芥的CERK1却能在本氏烟细胞中受flg22诱导进入警戒状态,提示通过基因编辑改造农作物自身CERK1的近膜区使之获得对应拟南芥CERK1的可磷酸化位点,有望成为提升农作物广谱真菌抗性而又不会造成持续生长抑制的新策略。
博士生龚本强为该论文的第一作者,李剑峰教授为通讯作者,前中大生科院的王宏斌教授参与了该研究。该研究获得国家自然科学基金的资助。
值得一提的是,2018年李剑峰和王宏斌教授作为共同通讯作者在Cell Host & Microbe发表了另一篇研究论文(10.1016/j.chom.2017.12.005),揭示了CERK1激酶区第428位酪氨酸磷酸化动态调控几丁质信号转导的机制。
参考文献
[1] Bahram M, et al. (2018). Structure and function of the global topsoil microbiome. Nature 560, 233-237.
[2] Fisher MC, et al. (2012). Emerging fungal threats to animal, plant and ecosystem health. Nature 484, 186-194.
[3] Chang Y, et al. (2015). Phylogenomic anlayses indicate that early fungi evolved digesting cell walls of algal ancestors of land plants. Genome Biol. Evol. 7, 1590-1601.
[4] Fisher MC, et al. (2018). Worldwide emergence of resistance to antifungal drugs challenges human health and food security. Science 360, 739-742.
[5] Bressendorff S, et al. (2016). An innate immunity pathway in the moss Physcomitrella patens. Plant Cell 28, 1328-1342.
论文链接:
https://www.cell.com/cell-host-microbe/fulltext/S1931-3128(19)30534-7
点评链接:
https://www.cell.com/cell-host-microbe/fulltext/S1931-3128(19)30589-X
来源:meta-genome 宏基因组
原文链接:https://mp.weixin.qq.com/s?__biz=MzUzMjA4Njc1MA==&mid=2247489255&idx=2&sn=75c4949eb87d224773f2d6cb0b79b25a&chksm=fab9fc56cdce7540fd3826c80fa564487be35eb67a3b5eba4abd760f0ddd158c5981c6c78b63#rd
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn

面对病菌,植物会开启“诱敌”模式

最新研究揭示转多基因赋予小麦对病原真菌的广谱抗性!
云南大学王建光、陈穗云研究团队揭示真菌多肽介导的TMV抗性机制

Plant Cell :真菌几丁质酶类效应子抑制植物免疫
Nature Plants:植物根部共栖微生物调节植物免疫反应,避免过度免疫

四川农大陈学伟组综述来非编码RNA在植物免疫过程中作用

发现第二种植物胞外ATP受体,揭示其在植物免疫中的作用
《Mycology》热点文章推送——曲霉抗真菌药物的抗性机制及作用蛋白
植物逆境中心Alberto Macho组揭示抑制植物免疫新机制
植物逆境中心揭示植物感受鞭毛的新型免疫受体


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号