

宫奥博
加好友
宫奥博 2021-10-11
作者:十一月

图1 细胞中的多种液-液相分离介导形成的凝聚体【1】
活细胞中所有结构的形成都需要一定的时序以及空间调控,这其中就包括诸多由生物大分子液-液相分离特性所介导形成的凝聚体【1】,比如核仁、Cajal小体以及应激颗粒等均是通过富含内在无序序列的蛋白质或核酸生物分子网络形成的(图1)。但是一直以来关于液-液相分离在生物体内的活性与其功能之间的因果关系以及定量化描述等方面还存在诸多争议(详见:特别推荐 | 相分离与其生理功能是否存在因果关系?)。
为了对活细胞中的大分子凝聚体成核作用进行定量化的模型分析,2021年9月22日,美国普林斯顿大学Clifford P. Brangwynne研究组发文题为Nucleation landscape of biomolecular condensates,引入了非活细胞体系中的经典成核理论(Classical nucleation theory,CNT)对活细胞内的液滴形成过程的成核速率以及成核位置的描述,给成核作用以更为数学模型化以及定量化的测量方式。

在非生命系统中,当热波动克服表面张力带来的能量壁垒时,初始阶段的相分离成核作用开始出现,这一现象可以用经典的成核理论进行模拟,该理论中提到液滴的成核速率依赖于过饱和的程度,而液滴出现的位置则是由稀释相与浓缩相界面的非均质的特征所控制的【2】。但是由于活细胞内环境中组分众多且高度复杂,存在多种不同的内在无序序列,经典的成核理论是否适用于活细胞尚不清楚。
为了量化活细胞中凝聚体成核作用,作者们测量了体内凝聚体的成核速率J,J的定义是单位体积、单位时间内凝聚体形成的数量。进一步地,为了对成核速率的决定因素进行探究,作者们引入了Corelets系统,Corelets是一种光遗传学工具,能够利用光在空间和时间上控制细胞内液相分离【3】。然后作者们将已知能够发生相分离的内在无序序列引入该系统之中,对成核作用的速率进行探究(图2)。

图2 利用Corelets系统对可以发生相分离的内在无序序列进行定量光遗传学控制
相分离凝聚体的形成包括两个过程,一是成核作用,另一个是生长过程。作者们对控制成核作用因素探究后,建立起了一个用以描述成核速率的方程(图3)。S*是一个无量纲参数,用以描述成核作用激活和运输限制的交叉过饱和程度。当细胞内生物分子的浓度低于过饱和浓度时,成核作用强烈地依赖于饱和浓度的激活;但是当浓度远远高于过饱和浓度时,成核作用就会受到微观运输作用限制以及凝聚体成核作用位点(多价相互作用位点)密度的限制。另外,由于S*也会受到表面张力的影响,而且生物大分子会在特定的位点进行凝聚,这说明在成核作用中的生物大分子会作为“种子”发挥作用【4,5】。因此,作者们将生物大分子凝聚体的表面张力以及“种子”作用的能力也纳入成核作用理论的参数考虑之内。为了对经典成核作用理论进行进一步地拓展,作者们建立了在特定位点引入相分离凝聚体的实验方法,将已知能够发生相分离的内在无序序列与端粒重复序列结合蛋白质TRF1构建成为融合体系并同时联用光遗传系统Corelets,从而可以在特定的端粒处对成核作用进行描述。进一步地,为了对S*决定因素进行进一步地细化,作者们对内在无序序列中介导其相互作用的关键氨基酸进行突变,发现参数S*依赖于生物大分子本身作为成核作用“种子”的能力,也就是内在无序序列之间相互作用的能力。

图3 成核速率方程
最后为了检测成核理论方程在内源生理以及异常条件下对凝聚体的成核过程是否能够进行描述,作者们使用Corelet系统对应激颗粒进行标记。通过对应激颗粒组装关键因子G3BP敲除的细胞中回补表达G3BP-Corelet系统以及加入诱发应激颗粒产生的砷酸盐,作者们确认砷酸盐诱导后应激颗粒产生的速度更快,但未处理的和加入砷酸盐的细胞中应激颗粒的产生均能落在方程曲线上,该结果说明了成核理论方程的适用性。
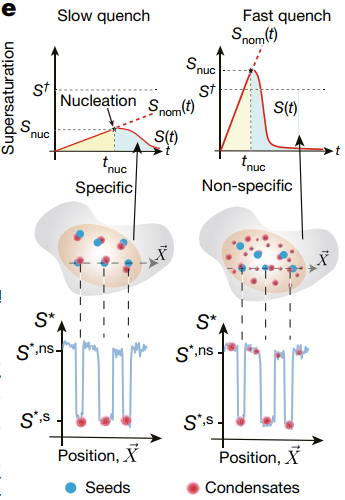
图4活细胞内成核作用景观示意图
总的来说,作者们提出了一个用以量化和和描述外源的以及内源生理水平生物大分子凝聚体的成核作用的数学模型(图4)。细胞内的环境非常复杂,但作者们得到的成核作用方程可以用来解释生物大分子成核速率对于过饱和浓度、内在无序序列之间的相互作用等方面的依赖。该框架有助于帮助理解各种生物大分子凝聚体的成核作用以及生长过程的动力学特征,为生物工程化液-液相分离凝聚体提供了新的参数辅助。
原文链接:
https://www.nature.com/articles/s41586-021-03905-5
参考文献
1 Shin, Y. & Brangwynne, C. P. Liquid phase condensation in cell physiology and disease. Science (New York, N.Y.) 357, doi:10.1126/science.aaf4382 (2017).
2 Auer, S. & Frenkel, D. Prediction of absolute crystal-nucleation rate in hard-sphere colloids. Nature 409, 1020-1023, doi:10.1038/35059035 (2001).
3 Bracha, D. et al. Mapping Local and Global Liquid Phase Behavior in Living Cells Using Photo-Oligomerizable Seeds. Cell 175, 1467-1480.e1413, doi:10.1016/j.cell.2018.10.048 (2018).
4 Shevtsov, S. P. & Dundr, M. Nucleation of nuclear bodies by RNA. Nature cell biology 13, 167-173, doi:10.1038/ncb2157 (2011).
5 Berry, J., Weber, S. C., Vaidya, N., Haataja, M. & Brangwynne, C. P. RNA transcription modulates phase transition-driven nuclear body assembly. Proceedings of the National Academy of Sciences of the United States of America 112, E5237-5245, doi:10.1073/pnas.1509317112 (2015).
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号