科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2020-07-04
来源:植物科学最前沿
小分子信号转导是植物对外界环境和内部信号的响应的重要机制。植物将大量输入信号整合成一个连贯的整体信号通路,广泛调控植物生长发育。
ARFs转录水平调控
ARFs可以分成3个家族:A-ARFs、B-ARFs和C-ARFs。通常认为A-ARFs是转录激活子,B-ARFs和C-ARFs是转录抑制子。虽然大多数A类ARF在根的分生组织中表达,但研究发现ARF8的表达仅限于外层细胞,而ARF19的表达仅限于根冠、表皮和中柱细胞中。因此,在任何类型的细胞中存在哪些ARF取决于细胞的发育阶段和定位。ARF表达的时空调控导致不同的ARF在不同组织中存在,对于生长素的响应也取决于这些组织中存在什么样其他因素。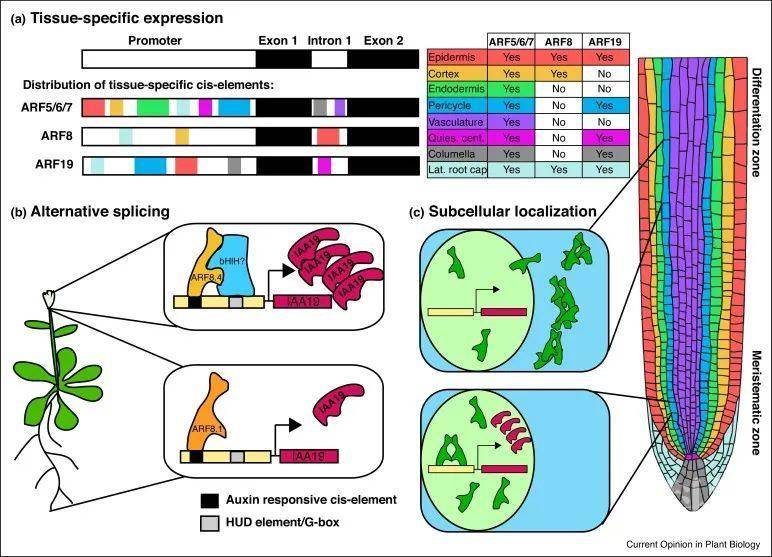
图1 在转录和蛋白水平上对AFR功能的调控
选择性剪接是调控ARF表达和相互作用另一种方法。近来研究表明ARF8剪接变体在表达谱上存在明显的差异,例如剪接变体8.4在发育中的拟南芥花序中特异表达。与两个全局性表达剪接变体ARF8.1和ARF8.2相比,8.4存在28个氨基酸插入。与ARF8.1和ARF8.2不同,在ARF8-7突变体中超表达ARF8.4能够完全恢复生长素响应基因IAA19的转录。研究发现,ARF8.4与IAA19启动子区域的结合强度高于其他变体,而这些区域中包含了能够供其他转录因子的结合的G-box和HUD等元件。因此推测ARF8.4可能在这个启动子上表现出其他变体没有的协同活性。同样地,在番茄中也观察到了ARF的剪接变体,这些变体在生殖组织中也表现出组织特异性的表达。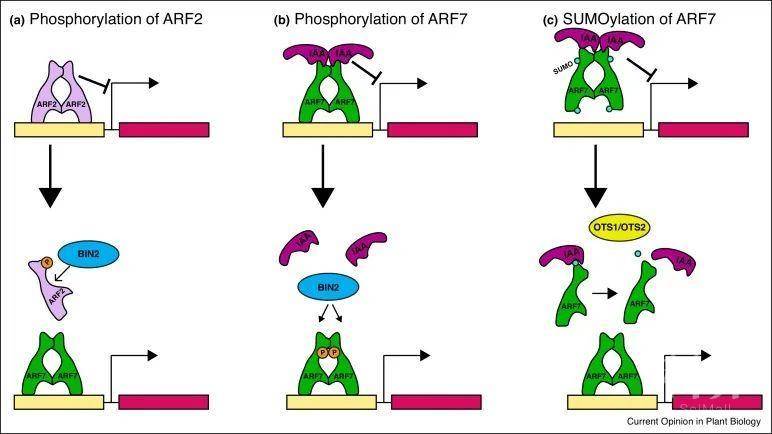
图2 ARF活性的翻译后调控
ARFs在蛋白结构和定位上的调控
转录因子在细胞核中发挥作用,因此蛋白定位是植物调节转录因子活性的另一个关键机制。最近研究表明,ARF7和ARF19定位在初生根分生区的细胞核内,但在根的分化区域却发生了重定位。除此之外,ARF的翻译后修饰也是发挥调控作用的重要方式,但这些修饰如何相互作用或促进协同性尚不清楚。对一些家族成员来说,ARF的磷酸化是ARF功能的关键。例如油菜素甾体调节激酶BIN2对B类ARF2的磷酸化降低了其与DNA的结合能力,并且因为ARF2是一种转录抑制因子,这就可能增强生长素的响应能力。BIN2也可以磷酸化A类ARF7和ARF19,干扰ARF与其IAA抑制剂的相互作用来促进ARF与DNA的结合和生长素响应。ARFs和其他转录因子的互作影响植物发育
转录的组合调控是信号通路之间协同作用的一个重要标志,即来自多个途径的转录因子与启动子间相互作用。尽管关于生长素响应如何受其他信号途径影响的研究很多,但有关ARFs与启动子上其他转录因子之间的相互作用的报道相对较少。对拟南芥生长素响应蛋白的自然变异分析发现,ARF的自然变异集中在几个家族成员的可变中间域和C端,而这些区域是蛋白质相互作用的区域,表明ARF蛋白的相互作用发生的范围很广,也可能是多样性进化的目标。转录的组合调控标记
启动子结构分析是寻找转录组合调控的另一个方向,最近对生长素响应启动子的研究表明,拟南芥生长素响应启动子上广泛存在转录因子共占有的情况。研究发现,对生长素反应迅速上调的基因的启动子可能是直接的ARF靶点,这些启动子富集着TCP、bZIP和bHLH转录因子类的结合位点。独立于转录调节的发育弹性
转录调控中的合作性产生了发育的复杂性,这种复杂性带来了系统内噪音的增加和阻碍连贯性反应的风险。由于植物必须在不断变化的环境中生长,对微小环境扰动的过度反应会对植物的适应性产生负面影响。因此,作为一种独立于转录调节的方法,植物可以利用生物力学约束来限制这种噪音。例如,最近研究发现,拟南芥液泡大小也可以独立于生长素调节,肌动结合蛋白NET4也可以促进其收缩。NET4对液泡大小的影响并不依赖于生长素的处理,这表明这种细胞骨架调节器提供了一种调节器官生长和组织扩张的正交机制,可以起到防止有害转录波动的作用。来源:frontiersin 植物科学最前沿
原文链接:https://mp.weixin.qq.com/s?__biz=MzIyOTY2NDYyNQ==&mid=2247497623&idx=3&sn=3c6c1ff91ce59f976bcb221fdd460e43&chksm=e8bd8589dfca0c9f74e93bf3bd5b3064363fb597ed24ae3a06c55f7500bb3bcf989cfbad1a1a#rd
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn

全面分析拟南芥、玉米和高粱核心启动子,助力设计新启动子和作物改良

Nature Plants | 揭秘最原始的生长素响应系统!

Nature 重磅| 逆境中心、福建农林等单位发现新的生长素信号途径

GmYUC2a是大豆结瘤过程中生长素生物合成的重要调控基因!

EMBO J | 生长素极性运输调控新机制
生工小课堂|启动子和终止子

林鸿宣研究组揭示局部生长素合成调控水稻株型和穗型发育的分子机理
首次发现DRO1基因启动子中的INDITTO2转座子可传递生长素信号

生长素如何指导根在向地生长时避开障碍?

生长素调控细胞生长的分子机理


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号