科技工作者之家
科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
科技工作者之家 2020-12-04
来源:ACS美国化学会
供稿:李佼峰,北京大学
大家好,今天为大家分享一篇ACS Chemical Biology的文章,题为“Addicting Escherichia coli to New-to-Nature Reactions”,本文的通讯作者为荷兰格罗宁根大学的Clemens Mayer教授。
对开放系统中的生物进行基因编辑时,人们需要格外关注生物安全问题,防止逃逸变异株对生态系统造成负面影响,因此必须采取严格有效的生物安全防护措施以限制其在自然环境下的增殖。在此前的工作中,非天然氨基酸依赖型的营养缺陷株已被用于解决这一潜在问题;但为了使其进一步远离生态系统,本文作者开发了新一代营养缺陷型大肠杆菌菌株,通过过渡金属催化的生物相容反应,原位生成非天然氨基酸以供其依赖型营养缺陷株生长。
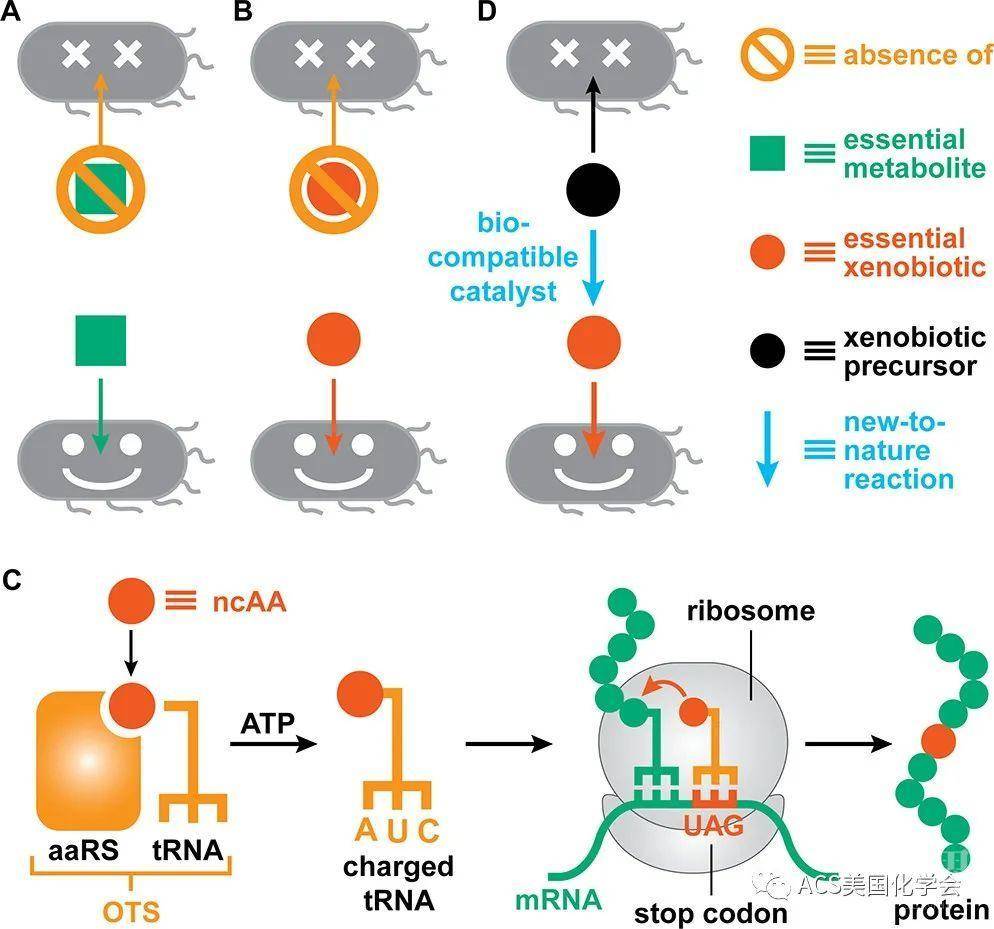
图1. 采用营养缺陷株进行生物防护。A.依赖必需代谢物的营养缺陷株;B.依赖外源物质(非天然氨基酸)的营养缺陷株;C.遗传密码子拓展技术将非天然氨基酸引入蛋白质;D.依赖生物相容催化剂原位形成非天然氨基酸的营养缺陷株。
为了创建细菌生存和非天然氨基酸之间的关联,作者将最近报道的β-内酰胺酶的变体TEM-1.B98引入大肠杆菌中。由于这一变体降解氨苄青霉素(A抗)的活性依赖于3iY(3-碘酪氨酸)或3nY(3-硝基酪氨酸)的插入,故该菌株在氨苄青霉素中的生长严格依赖于外源的3iY/3nY。首先,作者证明在A抗平板上,只有提供外源3iY/3nY,才能观测到菌落形成(如图2B)。且在液体培养基中,非天然氨基酸浓度提高使得细菌能够在较高抗生素浓度的条件下生长(如图2C)。其中由于3nY能够使得细菌在更高A抗浓度下生长,作者选用3nY进行后续实验。
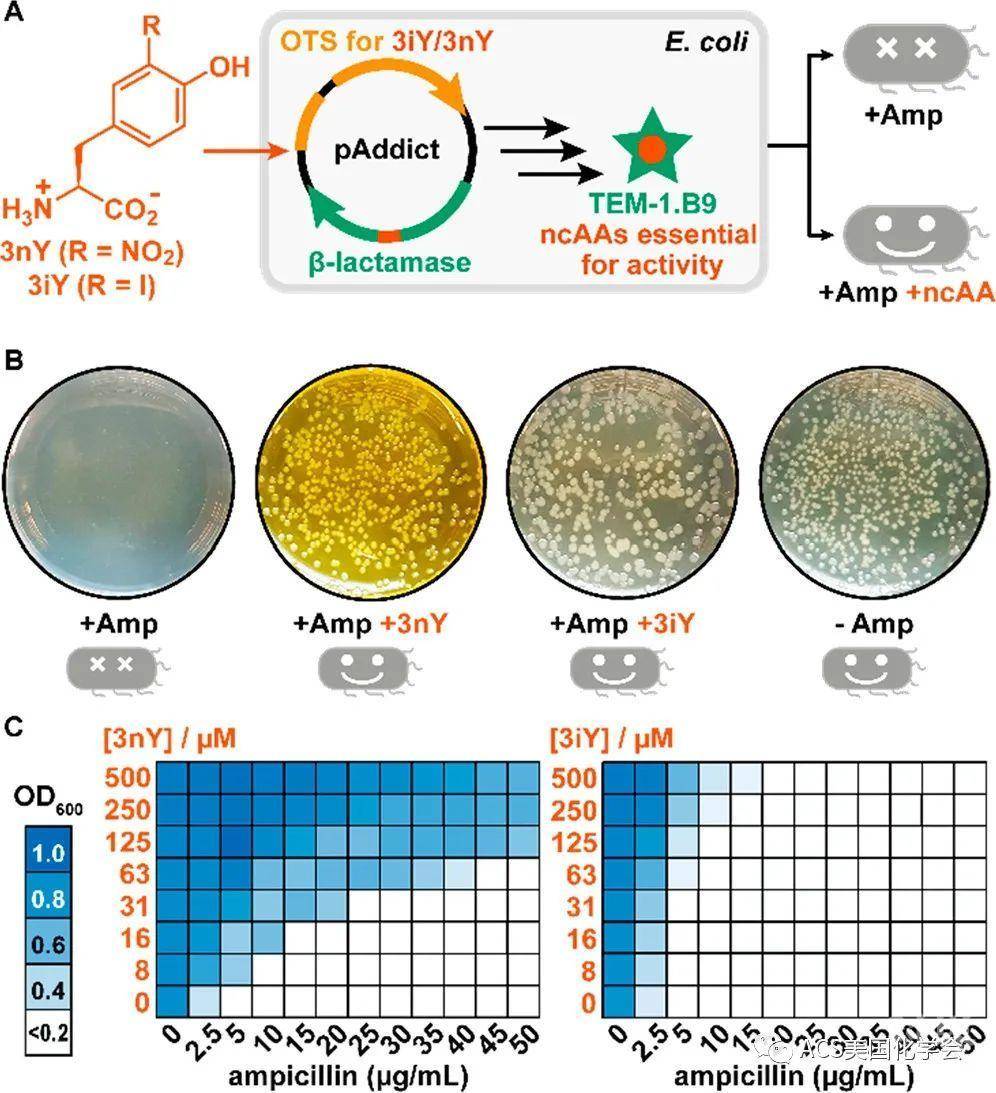
图2. 营养缺陷株依赖于非天然氨基酸供应而生存。A.依赖于非天然氨基酸的菌株质粒和生存示意图;B.营养缺陷株在抗生素和/或非天然氨基酸的平板上生长情况;C.营养缺陷株生长密度随抗生素和非天然氨基酸浓度改变。
为了利用非天然氨基酸的原位转化调控细菌生长,作者选择alloc-3nY(烯丙氧羰基保护3-硝基酪氨酸)作为非天然氨基酸前体,而催化剂则选用能够在大肠杆菌中循环利用且毒性较小的钌配合物 Ru1(如图3A、B)。为了评价alloc-3nY存在下,Ru1促使细菌生存的能力,作者检测不同A抗浓度下细菌生长的密度,发现alloc-3nY本身无法使细菌生存,而在Ru1催化下能够促进其生存。催化剂浓度增加使得细菌在更高抗生素浓度下得以生存,而当催化剂载量高于2.5%时,细菌增长的速度可以和仅提供3nY时相媲美。此外相比同时添加Ru1和3nY,Ru1和alloc-3nY共同处理会使得细菌在更高抗生素浓度下得以生长(如图3D)。作者猜测该现象是由于Ru1和3nY对细胞有一些毒性,而3nY的前体alloc-3nY无毒;因此相比直接提供3nY,Ru1催化3nY的持续产生更加有利于细菌生长。
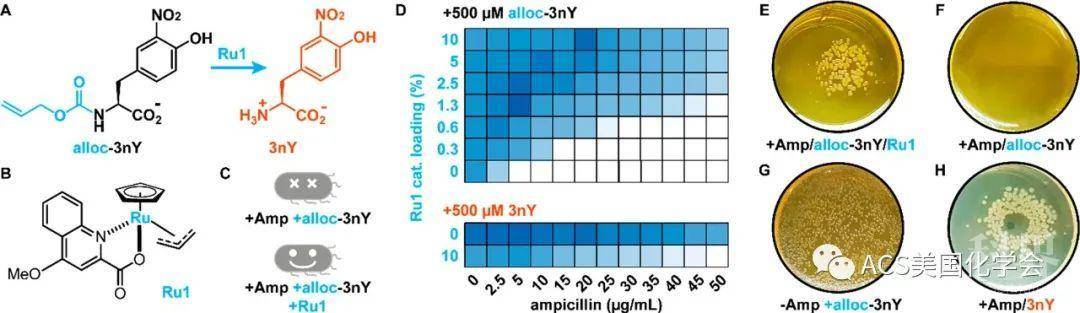
图3. 均相Ru催化剂挽救营养缺陷株生长。A. Ru催化的脱保护反应;B.催化剂结构;C.营养缺陷株生长示意图;D. 营养缺陷株分别被提供非天然氨基酸或前体及催化剂组合后,生长密度随抗生素浓度和催化剂载量变化;E/F/G/H. 营养缺陷株在抗生素、非天然氨基酸或其前体,催化剂存在的平板上生长情况。
随后,为检测Ru1是否能在固体培养基上促进细菌生长,作者向平板上添加Ru1储液后任其自由扩散,并检测菌落形成。在48-72 h后,作者发现仅在Ru1附近观测到菌落的产生(如图3E);而在只添加3nY以模拟Ru1扩散时,作者观察到光晕模式(如图3H,中间无菌生长)。这些结果表明:相比直接提供3nY,利用生物相容催化剂介导3nY原位的产生不会展现出明显毒性。
为了进一步调节细菌的生长且允许催化剂的回收利用,作者制备了固定在聚苯乙烯微球上的Pd纳米颗粒,将其用于炔丙基保护的氨基原位脱保护(如图4A)。作者合成了proc-p-ClF和proc-3nY作为非天然氨基酸前体,并以GFP Y151TAG突变体作为模型检测新鲜制备Pd纳米颗粒的生物活性。当proc-p-ClF作为底物,且Pd纳米颗粒浓度高于5 mg/ml时,作者观测到荧光信号的显著增长,证明了该反应的高效性。
最后作者评价proc-3nY在Pd纳米颗粒催化下挽救营养缺陷株生长的能力。在液体培养基中添加Pd纳米颗粒、proc-3nY和A抗时,细菌会在24 h后显著增长。为了证明细菌的生长是由Pd纳米颗粒催化氨基酸前体转换介导的,作者利用HPLC对3nY的浓度进行定量,发现非天然氨基酸的浓度与观测到的细胞密度呈正相关(如图4B)。最后作者测试催化剂是否可以重复回收利用,在每个循环中,他们定量检测细菌的生长密度和3nY浓度,发现第三次回收之后催化活性才会有所下降(如图4D),证明了异相金属催化剂可被重复利用以调控非天然氨基酸依赖型的营养缺陷株生长。
本文调控菌株生存的金属脱保护生物相容反应1也被称作金属催化生物正交剪切反应,已被广泛应用于小分子前药激活,蛋白质功能调控和细胞工程化等多个方面,主要包括钌2、钯3、金4介导的烯丙氧羰基,铜介导的二甲基取代炔丙氧羰基5和钯介导的炔丙基6脱保护反应。生物正交剪切反应最开始被用于活体中荧光小分子的激活;随后通过和遗传密码子拓展策略联用,科学家们实现了活细胞内蛋白质的特异激活7。而本文作者反其道而行之,先采用生物正交剪切反应催化非天然氨基酸的原位生成,后利用遗传密码子拓展策略调控非天然氨基酸依赖型营养缺陷株的生长,完全阻断了基因向野生型生物传播,进一步加固了野生生物和改造生物之间的屏障,在生物防护方面有明显优势。
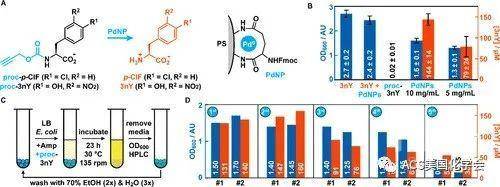
图4. 异相Pd催化剂挽救合成营养缺陷株。A. Pd催化的脱保护反应及催化剂结构;B.细菌密度、非天然氨基酸生成对催化剂和前体的依赖;C.多次回收利用Pd纳米颗粒;D.回收利用的Pd催化效率和细菌生长情况。
参考文献
1. Nat Chem Biol 12, 129-137, (2016).
2. Angew Chem Int Ed Engl 45, 5645-5648, (2006).
3. J Am Chem Soc 131, 5163-5171, (2009).
4. Angew Chem Int Ed Engl 56, 12548-12552, (2017).
5. Bioconjug Chem 24, 684-689, (2013).
6. Chem Commun (Camb) 46, 3964-3966, (2010).
7. Nat Chem 6, 352-361, (2014).
8. Nat Chem Biol 12, 138-140, (2016).
来源:gh_0320d0d498b4 ACS美国化学会
原文链接:http://mp.weixin.qq.com/s?__biz=MzIyMjgzMzI0Ng==&mid=2247508597&idx=1&sn=5458f218522ea32023b8be0f7e72f60b
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn

不只催化剂,助催化剂也要单原子化! | NSR研究论文
固体催化剂
能源催化剂

这种二维材料虽易自氧化,但或可作为高效催化剂

“智能穿梭”聚合物催化剂,助力ppm级Pd催化交叉偶联、及催化剂连续流动回收利用

新型制氢催化剂——离清洁能源更进一步
催化剂提高溶剂运动速率
钙钛矿催化剂

【视频】魔术神杯催化剂

冯小明:二十年磨一剑,耐得住寂寞的科学家 | 2018年未来科学大奖获奖人


科技工作者之家APP是专注科技人才,知识分享与人才交流的服务平台。
 微信
微信
 京公网安备11010202008424号
京公网安备11010202008424号